
 A capa de Fevereiro do PLoS Pathogens é dedicada a um velho conhecido nosso, o Toxoplasma gondii, um parasita que infecta uma grande parte da população humana. Num artigo (ref1) da PLoS Pathogens, Ke Hu e colegas estudaram em detalhe a máquina invasora por detrás do toxoplasma, o chamado Complexo Apical, , que é parte da estrutura que forma o cito-esqueleto do parasita. [... ler mais]
A capa de Fevereiro do PLoS Pathogens é dedicada a um velho conhecido nosso, o Toxoplasma gondii, um parasita que infecta uma grande parte da população humana. Num artigo (ref1) da PLoS Pathogens, Ke Hu e colegas estudaram em detalhe a máquina invasora por detrás do toxoplasma, o chamado Complexo Apical, , que é parte da estrutura que forma o cito-esqueleto do parasita. [... ler mais]
Contrariamente à ideia de um organismo unicelular como uma espécie de bolha amorfa, o toxoplasma possui uma espécie de esqueleto com extremidades bem definidas, e crê-se que essa estrutura desempenha um papel importante, de tipo mecânico, na invasão das células do hospedeiro pelo parasita.Os detalhes do artigo são algo técnicos mas a motivação dos autores é simples: uma melhor compreensão de como o complexo apical faz o seu trabalho poderá levar a novas ideias para combater as doenças criadas por esta família de parasitas.
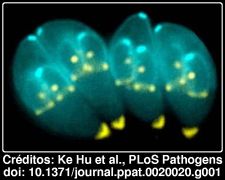
A máquina molecular por trás deste complexo era desconhecida mas Ke Hu e colegas desenvolveram técnicas que lhes permitiram identificar a maior parte das proteínas usadas para construir o complexo apical. As imagens obtidas pelos autores são fantásticas. Na imagem acima, onde as diferentes cores correspondem a diferentes proteínas, os processos utlizados pelos autores permitem-nos mesmo ver pequenas células-filha dentro das células-mãe. Ou seja, os autores poderam mesmo seguir a génese desta estrutura. Numa tradução livre do resumo:Acredita-se que o complexo apical do Toxoplasma gondii desempenha funções essenciais quer na invasão das células hospedeiro (incluindo células humanas), quer na replicação do parasita. Compreender a função do complexo apical, a base para esta estrutura inovadora, e o mecanismo para a sua motilidade, são dificultados pela falta de conhecimento da sua composição molecular. Nós purificámos parcialmente o complexo conoidal/apical, identificámos ~200 proteínas que representam 70% dos componentes proteicos cito-esqueléticos, caracterizámos sete novas proteínas, e determinámos a sequência de recrutamento de cinco dessas proteínas no cito-esqueleto durante a divisão celular. Os nossos resultados fornecem novos marcadores para os diferentes subcompartimentos dentro do complexo apical, e revelaram compartimentos celulares desconhecidos anteriormente, que facilitam a nossa compreensão de como a maquinaria da invasão é construída. Surpreendentemente, as estruturas nos extremos apicais e basais desta célula altamente polarizada originam-se no mesmo local e ao mesmo tempo durante os estágios inicias da replicação do parasita.
A importância deste tipo de estudos não se esgota neste parasita, o Toxoplasma gondii é apenas uma entre cerca de 5,000 espécies de protozoários parasitas intracelulares no filo Apicomplexa, que inclui outro velho conhecido nosso, o agente causador da malária, o Plasmodium falciparum. Uma das gandes vantagens das revistas da Public Library of Science (PLoS) é que são completamente livres, todo o material poder ser disseminado sem preocupações de direitos de autor. Não resisto por isso a mostrar mais uma das fantásticas imagens do artigo, em que mais uma se vez se veêm toxoplasmas dentro de toxoplasmas. As dimensões dos pormenores são ínfimas, 1μ corresponde a um milésimo de milímetro.
Referências
(ref1) Hu K, Johnson J, Florens L, Fraunholz M, Suravajjala S, et al. (2006). Cytoskeletal Components of an Invasion Machine: The Apical Complex of Toxoplasma gondii. PLoS Pathog 2(2): e13. Laço DOI
terça-feira, fevereiro 28, 2006
Visão nova de um cliente antigo
segunda-feira, fevereiro 27, 2006
Não apenas deliciosa mas também bonita

 A mais nobre das criaturas aquáticas, a ilustre lampreia, não é apenas uma iguaria gastronómica sem rival, é também importante pelo vislumbre que nos fornece da história dos vertebrados. Os "peixes verdadeiros" possuem uma boca com mandíbulas, mas os antepassados dos peixes actuais, até há cerca de 500 milhões de anos atrás, não eram capazes de morder, e as lampreias mostram ainda essa condição ancestral. As lampreias apresentam como boca uma espécie de ventosa coberta coberta de dentes, com que se agarram à carne dos peixes, aos quais chupam o sangue. As diferenças das lampreias em relação aos peixes não se esgotam na boca, existe um vasto reportório de características fisiológicas e morfológicas que separam ambas as categorias de vertebrados. Entre algumas peculiaridades da lampreia estão um sistema imunitário completamente diferente do nosso, e uma grande tolerância ao ferro. [... ler mais]
A mais nobre das criaturas aquáticas, a ilustre lampreia, não é apenas uma iguaria gastronómica sem rival, é também importante pelo vislumbre que nos fornece da história dos vertebrados. Os "peixes verdadeiros" possuem uma boca com mandíbulas, mas os antepassados dos peixes actuais, até há cerca de 500 milhões de anos atrás, não eram capazes de morder, e as lampreias mostram ainda essa condição ancestral. As lampreias apresentam como boca uma espécie de ventosa coberta coberta de dentes, com que se agarram à carne dos peixes, aos quais chupam o sangue. As diferenças das lampreias em relação aos peixes não se esgotam na boca, existe um vasto reportório de características fisiológicas e morfológicas que separam ambas as categorias de vertebrados. Entre algumas peculiaridades da lampreia estão um sistema imunitário completamente diferente do nosso, e uma grande tolerância ao ferro. [... ler mais] As lampreias não são o único grupo de peixes sem mandíbulas, as mixinas ou enguias de casulo também não as possuem. Em vez de maxilares que se articulam verticalmente como os nossos, as mixinas possuem duas estruturas que se articulam horizontalmente para puxarem comida. Uma imagem destas simpáticas criaturas, caracterizadas por tentáculos próximos da boca, mostra-se aqui ao lado. Originalmente, as mixinas e as lampreias eram incluídas num grupo designado Cyclostomi (ciclóstomos). Contudo, há relativamente pouco tempo, alguns investigadores favoreceram a hipótese de que as lampreias possuíam um antepassado comum mais recente em relação aos peixes com mandíbulas, os chamados Gnathostomata (gnatóstomos), que em relação às mixinas. A relação entre os três grupos mantêm-se no entanto algo incerta pois trabalhos recentes com ADN mitocondrial (ref2) sugerem que as mixinas e as lampreias estão afinal mais próximas umas das outras que dos gnatóstomos, e a tendência actual é incluir quer as mixinas quer as lampreias nos vertebrados.
As lampreias não são o único grupo de peixes sem mandíbulas, as mixinas ou enguias de casulo também não as possuem. Em vez de maxilares que se articulam verticalmente como os nossos, as mixinas possuem duas estruturas que se articulam horizontalmente para puxarem comida. Uma imagem destas simpáticas criaturas, caracterizadas por tentáculos próximos da boca, mostra-se aqui ao lado. Originalmente, as mixinas e as lampreias eram incluídas num grupo designado Cyclostomi (ciclóstomos). Contudo, há relativamente pouco tempo, alguns investigadores favoreceram a hipótese de que as lampreias possuíam um antepassado comum mais recente em relação aos peixes com mandíbulas, os chamados Gnathostomata (gnatóstomos), que em relação às mixinas. A relação entre os três grupos mantêm-se no entanto algo incerta pois trabalhos recentes com ADN mitocondrial (ref2) sugerem que as mixinas e as lampreias estão afinal mais próximas umas das outras que dos gnatóstomos, e a tendência actual é incluir quer as mixinas quer as lampreias nos vertebrados.
As lampreias possuem numa das suas extremidades uma cabeça sustentada por cartilagem, e não possuem ossos. Grande parte dos problemas em colocar a lampreia entre os vertebrados tinha a ver com as supostas peculiaridades do esqueleto da lampreia, que se supunha ser bastante diferente do dos restantes vertebrados e ter evoluído separadamente. Em particular supunha-se que a cartilagem da lampreia era diferente da cartilagem dos restantes vertebrados que se baseia em colagénio. Quando se fala em colagénio, a maioria das pessoas pensa imediatamente nos lábios das estrelas de cinema mas o colagénio tem um papel vital no desenvolvimento do esqueleto dos seres humanos. O colagénio é uma estrutura proteica longa e fina que se encontra sobre a forma de feixes de fibras brancas, que apesar de flexíveis são muito resistentes à tracção. Na verdade a maioria das pessoas conhece muito bem o colagénio, pois quando aquecidas a temperaturas elevadas e arrefecidas em seguida estas fibras transformam-se em gelatina. Uma variante do colagénio, o colagénio de tipo II, é nos seres humanos e restantes vertebrados com mandíbulas, a fibra da cartilagem, que é não só um tecido de sustentação em si mesma mas é também uma etapa do crescimento do tecido ósseo.
Pensava-se que o colagénio de tipo II era uma invenção relativamente recente que unia grupos de vertebrados como répteis, anfíbios, tubarões e peixes com esqueleto ósseo, e os separava de vertebrados mais "primitivos" como a lampreia, cujo esqueleto seria baseado numa proteína diferente. De acordo com um artigo (ref1) publicado por GuangJun Zhang e colegas no Proceedings of the National Academy of Sciences as coisas não são bem assim. Apesar das centenas de milhões de anos de evolução que separam a lampreia da quase totalidade dos outros vertebrados, os genes que governam o desenvolvimento do seu esqueleto são os mesmos. Numa tradução livre do resumo:O colagénio de tipo II é a mais importante proteína da matriz da cartilagem no esqueleto dos vertebrados com mandíbulas. Pelo contrário, pensava-se que as lampreias e as mixinas possuíam uma cartilagem não-colagénica. Esta diferença na estrutura do esqueleto levou à hipótese que o antepassado comum aos vertebrados tinha um esqueleto não colagénico, com o colagénio de tipo II tornando-se a proteína dominante da matriz da cartilagem após a divergência entre os peixes sem mandíbulas dos peixes com mandíbulas há aproximadamente 500 milhões de anos atrás. Aqui mostramos que as lampreias têm dois genes de colagénio de tipo II (Col2α1) que se expressam durante o desenvolvimento do esqueleto cartilagíneo. Mostramos também que o esqueleto da lampreia adulta é rico na proteína Col2α1. Para além disso, isolámos um ortólogo da lampreia do Sox9, um regulador directo transcripcional de Col2α1 nos vertebrados com mandíbulas, e mostramos que se expressa com ambos os genes Col2α1 durante o desenvolvimento do esqueleto. Estes resultados mostram que o percurso genético para a condrogénese em lampreias e gnatóstomos é conservado durante a activação das moléculas da matriz cartilagínea e sugerem que um esqueleto com colagénio evoluiu supreendentemente cedo na evolução dos vertebrados.
Ou seja, a equipa de investigação usou técnicas de sequenciação de genes para mostrar que dois genes do colagénio de tipo II se expressam durante o desenvolvimento do esqueleto cartilaginoso da lampreia. Conseguiram ainda isolar um gene chamado Sox9, que regula a actividade do genes do colagénio nos gnastótomos. As implicações são simples: um esqueleto baseado no colagénio evoluiu antes do aparecimento dos gnastótomos, e antes que os antepassados das lampreias se tivessem separado destes. O resultado pode não parecer tão importante como isso mas é. Este estudo mostra que a característica comum dos vertebrados não é a presença ou ausência de tecido ósseo, mas sim partilharem um conjunto de genes que produz cartilagem.
Para finalizar nada melhor que uma imagem dos arquivos da EPA mostrando uma pobre truta a ser atacada por duas lampreias famintas.
Referências:
(ref1) GuangJun Zhang, Michael M. Miyamoto, and Martin J. Cohn (2006). Lamprey type II collagen and Sox9 reveal an ancient origin of the vertebrate collagenous skeleton. Proc. Natl. Acad. Sci. USA. doi: 10.1073/pnas.0508313103
(ref2) Delarbre et al (2002). Complete Mitochondrial DNA of the Hagfish, Eptatretus burgeri: The Comparative Analysis of Mitochondrial DNA Sequences Strongly Supports the Cyclostome Monophyly. Molecular Phylogenetics and Evolution 22 (2): 184-192.
Adenda: PZ Myers tem uma contribuição sobre este tema (em inglês), com imagens do artigo original, algo que eu em geral evito pois excepto pela PLoS biology a maioria das publicações não são livres e levam muito a sério os direitos de autor.
sábado, fevereiro 25, 2006
Ataque das galinhas mutantes de dentes afiados

 O Gallus gallus domesticus, mais conhecido pelas designações de galo, galinha ou frango tem sido presença assídua nos noticiários dos últimos tempos devido à famigerada gripe das aves. A ave mais comum à face da Terra, com mais de 20 mil milhões de exemplares, é para muitas pessoas sinónimo de alimento, sendo consumida em quantidades apreciáveis como frango assado, frito, na púcara, entre outras iguarias culinárias. Até recentemente ninguém associaria as galinhas a algo de sinistro, em parte porque a falta de dentes coloca as aves em desvantagem em relação aos seus primos próximos, os crocodilos, no que refere ao imaginário de criaturas assustadoras. Aliás é até vulgar associar um acontecimento invulgar a algo que sucederá no "dia em que as galinhas criem dentes". Pois bem, esse dia já chegou. [... ler mais]
O Gallus gallus domesticus, mais conhecido pelas designações de galo, galinha ou frango tem sido presença assídua nos noticiários dos últimos tempos devido à famigerada gripe das aves. A ave mais comum à face da Terra, com mais de 20 mil milhões de exemplares, é para muitas pessoas sinónimo de alimento, sendo consumida em quantidades apreciáveis como frango assado, frito, na púcara, entre outras iguarias culinárias. Até recentemente ninguém associaria as galinhas a algo de sinistro, em parte porque a falta de dentes coloca as aves em desvantagem em relação aos seus primos próximos, os crocodilos, no que refere ao imaginário de criaturas assustadoras. Aliás é até vulgar associar um acontecimento invulgar a algo que sucederá no "dia em que as galinhas criem dentes". Pois bem, esse dia já chegou. [... ler mais]
Num artigo na Current Biology (ref1), intitulado "The Development of Archosaurian First-Generation Teeth in a Chicken Mutant", Matthew Harris e colegas encontraram pequenos dentes num embrião com 16 dias. Numa tradução livre do resumo:As aves modernas não têm dentes. Em vez disso, desenvolveram uma estrutura queratinizada especializada, chamada a ranfoteca, que cobre a mandíbula, maxila e pré-maxila.
Os antepassados das aves actuais trocaram os dentes por um bico há qualquer coisa como 70 a 80 milhões de anos, e há mais de 65 milhões de anos que se extinguiram as últimas aves com dentes.Embora estudos de recombinação tenham mostrado que a epiderme aviana pode responder a sinais que induzam dentes a partir de mesênquima oral de ratos ou lagartos, as tentativas para iniciar de novo a formação de dentes em aves falharam.
Os autores referem aqui estudos dos anos 1980 em que se utilizou material retirado de tecido das gengivas de embriões de lagartos ou ratos para levar as galinhas a produzirem dentes. Contudo, o facto de se ter utilizado tecidos de outros animais não permitiu determinar até que ponto as aves mantinham o potencial para desenvolver de novo dentes, sobretudo porque por exemplo os dentes obtidos no caso da implantação de tecido de ratos se assemelhavam a dentes de ratos e não a dentes de fosséis de antepassados das aves actuais.Descrevemos aqui a formação de dentes numa galinha mutante talpid2, incluindo os processos de desenvolvimento e as mudanças moleculares iniciais associadas à formação de dentes. [...] Comparamos a formação de dentes no mutante talpid2 com o que acontece no aligator e mostramos a formação de dentes decididamente arcossaurianos (crocodilianos) de primeira geração no embrião da ave.
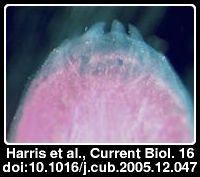 O talpid2 é um gene envolvido na formação de vários orgãos nas galinhas. Os dentes que se mostram na imagem ao lado são apenas três, e relativamente minúsculos, cobrindo apenas a extremidade da mandíbula. O que aconteceu é que nas aves não mutantes o gene que regula a formação dos dentes nos vertebrados se expressa no lado das gengivas, enquanto nas aves mutantes e nos crocodilos o gene se expressa nas células no topo das gengivas. Os dentes são cónicos e em forma de sabre, tal como os dos crocodilos e bastante diferente dos dentes dos mamíferos. Embora rara a mutação ocorre ocasionalmente mas acarreta problemas vários e os embriões morrem em geral antes dos 12 dias. Só num embrião que sobreviveu 16 dias (e que morreu há 50 anos) os cientistas se aperceberam que se estavam a formar dentes.
O talpid2 é um gene envolvido na formação de vários orgãos nas galinhas. Os dentes que se mostram na imagem ao lado são apenas três, e relativamente minúsculos, cobrindo apenas a extremidade da mandíbula. O que aconteceu é que nas aves não mutantes o gene que regula a formação dos dentes nos vertebrados se expressa no lado das gengivas, enquanto nas aves mutantes e nos crocodilos o gene se expressa nas células no topo das gengivas. Os dentes são cónicos e em forma de sabre, tal como os dos crocodilos e bastante diferente dos dentes dos mamíferos. Embora rara a mutação ocorre ocasionalmente mas acarreta problemas vários e os embriões morrem em geral antes dos 12 dias. Só num embrião que sobreviveu 16 dias (e que morreu há 50 anos) os cientistas se aperceberam que se estavam a formar dentes.
A semelhança com os dentes dos embriões dos crocodilos não é surpreendente, afinal de todos os grupos de animais existentes os crocodilos são os parentes mais próximos das aves. Tal como no caso dos peixes cegos que vivem em grutas discutido aqui algum tempo atrás, a perda ou atrofia de uma estrutura num organismo vivo não é um processo simples e a evolução segue por vezes caminhos tortuosos.
Referências
(ref1) The Development of Archosaurian First-Generation Teeth in a Chicken Mutant. Matthew P. Harris, Sean M. Hasso, Mark W.J. Ferguson, and John F. Fallon (2006). Current Biology, Vol 16, 371-377, 21. doi:10.1016/j.cub.2005.12.047
quarta-feira, fevereiro 15, 2006
Quando os necrófagos morrem

 Algo vai muito mal num ecossistema quando os organismos que se conseguem alimentar de corpos em decomposição desatam a morrer. Nos últimos dez anos os abutres iniciaram o caminho para o que se afigura a sua extinção no sub-continente indiano. As três espécies de abutre existentes na Índia, Gyps bengalensis, G. indicus e G. tenuirostris, sofreram uma derrocada populacional desde os anos 90 que atingiu índices alarmantes. Estima-se que o número de membros destas espécies tenha caido mais de 95%. Estamos a falar do desaparecimento de dezenas de milhões de aves. [... ler mais]
Algo vai muito mal num ecossistema quando os organismos que se conseguem alimentar de corpos em decomposição desatam a morrer. Nos últimos dez anos os abutres iniciaram o caminho para o que se afigura a sua extinção no sub-continente indiano. As três espécies de abutre existentes na Índia, Gyps bengalensis, G. indicus e G. tenuirostris, sofreram uma derrocada populacional desde os anos 90 que atingiu índices alarmantes. Estima-se que o número de membros destas espécies tenha caido mais de 95%. Estamos a falar do desaparecimento de dezenas de milhões de aves. [... ler mais]
Ao contrário da Europa, onde a perseguição por seres humanos erradicou as populações de grifos e abutres, na Índia as espécies sempre foram toleradas e protegidas, e o animal sempre foi valorizado por vários grupos religiosos. Por exemplo, os Parsis, de religião zaroastriana, não enterram os seus mortos, mas abandonam-nos para serem devorados pelos abutres, e assim não profanarem o solo, a água ou o fogo com um corpo impuro.
A dimensão da catástrofe deixou os investigadores inicialmente perplexos. Os animais mortos apresentavam lesões internas consistentes com um falhanço generalizado dos rins. Após uma série de testes concluiu-se que não se tratava de uma doença infecciosa e pensou-se imediatamente num contaminante ambiental. Só em 2004 se descobriu o culpado (ref1, ref2, ref3). Num exemplo de como as decisões humanas podem ter um impacto ecológico tremendo concluiu-se que a quase extinção dos abutres era devido a um anti-inflamatório utlizado para tratar os búfalos e vacas na Índia e nos estados vizinhos, o diclofenac. O que ninguém antecipou foi que, ao alimentar-se das carcaças dos animais mortos, os abutres fossem tão sensíveis aos efeitos da droga, e que uma tal mortandande fosse possível. A situação tornou-se crítica com os abutres oficialmente classificados como em risco de extinção. O desastre ecológico e sobre a saúde humana pode vir a ser considerável, com a explosão das populações de cães selvagens, e ratos, e com possibilidade de um aumento na transmissão de doenças como a raiva e a peste bubónica.
O governo da União Indiana anunciou em Março de 2005 a sua intenção da banir o produto do mercado indiano por volta de Setembro de 2005, mas a decisão esbarrou na falta de uma alternativa para o diclofenac. As perspectivas parecem finalmente ter melhorado com a publicação na PLoS Biology de um estudo de Swan e colegas (ref4) que mostrou que um outro medicamento, o meloxicam pode ser utilizado no gado sem efeitos nefastos sobre as populações de abutres. Para mais informação (em inglês) consultar as páginas da Darwin Initiative, Vulture Rescue, e BirdLife International. Este é mais um exemplo de como não considerar as consequências de impacto ambiental pode ter um efeito desastroso sobre um ecossistema. Ao introduzir um produto para o gado não basta pensar em como os animais domésticos e os seres humanos vão ser afectados, as consequências sobre todas as outras criaturas no ecossistema também têm que ser consideradas, em particular os animais no topo da cadeia alimentar como predadores e necrófagos.
Referências
(ref1) Oaks JL, Gilbert M, Virani MZ, Watson RT, Meteyer CU, et al. (2004) Diclofenac residues as the cause of vulture population declines in Pakistan. Nature 427: 630–633. versão online
(ref2) Shultz S, Baral HS, Charman S, Cunningham AA, Das D et al. (2004) Diclofenac poisoning is widespread in declining vulture populations across the Indian subcontinent. Proc R Soc Lond B Biol Sci (Suppl) 271: S458–S460 DOI: 10.1098/rsbl.2004.0223.
(ref3) Green RE, Newton I, Shultz S, Cunningham AA, Gilbert M, et al. (2004) Diclofenac poisoning as a cause of vulture population declines across the Indian subcontinent. J App Ecol 41: 793–800. versão online
(ref4) Swan G, Naidoo V, Cuthbert R, Green RE, Pain DJ, et al. (2006) Removing the Threat of Diclofenac to Critically Endangered Asian Vultures. PLoS Biol 4(3): e66. DOI: 10.1371/journal.pbio.0040066. versão online
O travo amargo do adoçante

 Muitos produtos utilizados na dieta alimentar das populações em países industrializados contêm um sem número de adoçantes artificiais, corantes, conservantes, aromas e sabores. Quando se junta a isso os processos de fabrico, embalagem, esterilização e distribuição de alimentos, os consumidores deveriam mostrar-se bastante mais apreensivos, sobretudo quanto a efeitos de longo prazo sobre a saúde, principalmente os aspectos cancerígenos. Claro que se pode sempre argumentar que existem inúmeros estudos do impacto destas substâncias sobre a saúde humana, e que os produtos e processos são seguros. Ou será que não? [... ler mais]
Muitos produtos utilizados na dieta alimentar das populações em países industrializados contêm um sem número de adoçantes artificiais, corantes, conservantes, aromas e sabores. Quando se junta a isso os processos de fabrico, embalagem, esterilização e distribuição de alimentos, os consumidores deveriam mostrar-se bastante mais apreensivos, sobretudo quanto a efeitos de longo prazo sobre a saúde, principalmente os aspectos cancerígenos. Claro que se pode sempre argumentar que existem inúmeros estudos do impacto destas substâncias sobre a saúde humana, e que os produtos e processos são seguros. Ou será que não? [... ler mais]
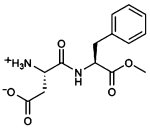
Num artigo na revista Environmental Health Perspectives, Morando Soffritti e colegas avaliaram o efeito do adoçante artificial aspartame (APM). Este produto é utilizado há mais de 30 anos como aditivo alimentar, sendo encontrado em mais de 6,000 produtos que incluem refrigerantes gaseificados, doces, pastilha elástica, iogurtes, e substitutos do açucar. Segundo o Centro de Informação do Aspartame (2005) o APM é consumido por 200 milhões de pessoas em todo o mundo. Sendo um produto tão largamente utilizado foi obviamente alvo de um vasto número de estudos. Morando Soffritti e colegas justificam no entanto a necessidade do estudo que levaram a cabo (tradução livre do original):Embora todos os estudos mencionados tenham sido considerados negativos no que respeita ao carácter carcinogénico do APM, na nossa opinião esses estudos não seguiram as regras básicas que hoje em dia se seguem para testar o potencial carcinogénico de um agente químico ou físico, em particular no que se refere ao número de animais em cada grupo experimental e à duração da experiência até às 110 semanas de idade dos animais.
A mega-experiência mencionada pelos autores durou cerca de 3 anos e consistiu em adicionar à dieta de uma amostra de 1,800 ratos de laboratório, divididos em diversos grupos de controlo, diferentes doses de APM. Os ratos foram então seguidos até ao fim das respectivas vidas (o último animal morreu com 159 semanas) e depois dissecados para avaliar os efeitos do APM. Os resultados são preocupantes, e eu vou poupar aqui a descrição da incidência de vários tipos de cancro que apareceram nos animais a quem foi ministrado APM quando comparados com um grupo aos quais não foi dada a substância. Digamos apenas que não deixam qualquer tipo de dúvidas. Passando directamente às conclusões:
Por estas razões, e à luz da difusão sempre crescente do APM na dieta dos países industrializados (em particular nos produtos consumidos por crianças pequenas e mulheres grávidas), considerámos ser importante realizar uma mega-experiência seguindo as boas regras laboratoriais, tal como são entendidas internacionalmente hoje em dia, para testes de carácter carcinogénico e, mais especificamente, os testes englobando todo o tempo de vida seguidos há já muitos anos no CMRC e descritos em publicações anteriores.O nosso estudo mostra que o APM é um composto carcinogénico com múltiplas potencialidades cujos efeitos são evidentes mesmo a doses diárias de 20 mg/kg de peso corporal (pc), muito menos que a dose diária indicada para seres humanos na Europa (40 mg/kg pc) e nos EUA (50 mg/kg pc).
Os resultados dos testes de carcinogenicidade em roedores são indicadores consistentes de riscos de cancro em seres humanos. Os resultados no nosso estudo chamam por isso por uma re-exanimação urgente das directivas actuais quanto ao uso e consumo de APM. A decisão de utilizar dados experimentais para protegar a saúde pública é importante porque o intervalo de tempo de uso alargado de APM é ainda demasiado breve para ter produzido evidência epidemiológica sólida. Para além disso, é pouco provável que dados suficientes se tornem disponíveis no futuro próximo, dada a dificuldade em encontrar um grupo de controlo que não tenha sido exposto a este composto amplamente difundido.
É óbvio que dado o peso económico do APM vai haver outros estudos, e que a palavra final sobre este tema ainda não foi dita. De facto, aquando de uma conferência em Setembro, em que foram apresentados resultados preliminares deste estudo, os fabricantes de APM questionaram a validade do estudo, que segundo eles entrava em conflito com centenas de estudos credíveis, que tenha sido revistos pelas entidades reguladoras em todo o mundo, e que as alegações eram inconsistentes com dados epidemiológicos humanos. Questionaram mesmo o passado do instituto responsável pelo estudo, tendo afirmado ser "criminoso" apresentar os dados publicamente antes de terem sido apresentados às entidades reguladoras, e antes de terem sido totalmente revistos. Enfim, o aspartame representa cerca de 62% do mercado de adoçantes artificiais, e a reacção, agora que o estudo foi publicado formalmente, vai ser provavelmente muito mais violenta. Mas para já é clara matéria para reflexão dos responsáveis políticos.
Referências
(ref1) Morando Soffritti, Fiorella Belpoggi, Davide Degli Esposti, Luca Lambertini, Eva Tibaldi, and Anna Rigano (2006). First Experimental Demonstration of the Multipotential Carcinogenic Effects of Aspartame Administered in the Feed to Sprague-Dawley Rats. Environ Health Perspect 114:379-385. Laço DOI.
terça-feira, fevereiro 14, 2006
De volta aos peixes das grutas
 Tal como prometido volto a focar a minha atenção no Astyanax mexicanus, o pequeno peixe que para além de uma forma de superfície com grandes olhos possui pelo menos trinta formas cegas que habitam uma série de grutas. [... ler mais]
Tal como prometido volto a focar a minha atenção no Astyanax mexicanus, o pequeno peixe que para além de uma forma de superfície com grandes olhos possui pelo menos trinta formas cegas que habitam uma série de grutas. [... ler mais]
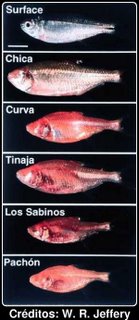
A imagem ao lado mostra algumas das variantes (Chica, Curva, etc. são os nomes das grutas).O Astyanax é um exemplo claro de evolução, no entanto alguns detractores da teoria da evolução das espécies por selecção natural apresentam-no como evidência contrária à evolução das espécies por modificação a partir das outras. Basicamente, o argumento, essencialmente da autoria de grupos religiosos, é que, embora desde a "expulsão do paraíso" as espécies estejam sujeitas a mutação e mudança, essas mudanças são em geral degenerativas e não conduzem ao aparecimento de estruturas complexas. Trata-se de um argumento recorrente que afirma que embora exista microevolução, em geral degenerativa e incapaz de produzir estruturas complexas, a macroevolução (aparecimento de novas espécies a partir de outras) não é possível.
Na verdade quando se analisa o Astyanax vê-se que as coisas não são assim tão simples como isso. Este peixe não se trata de uma versão corrompida da forma de superfície, nos aspectos que se revelam importantes para a vida numa gruta é mesmo uma versão melhorada. Por um lado, tal como se discutiu numa contribuição anterior, a degenerescência do olho é programada de forma complexa durante o desenvolvimento embrionário do peixe, e por outro lado o peixe compensa essa perda com o aperfeiçoamento de outros dos seus sentidos. De facto um artigo recente de Tamara Franz-Odendaal e Brian Hall (ref1) no Evolution & Development revê a forma como os diferentes módulos sensoriais da sardina de Mexico mostram diferentes tipos de adaptação à vida numa gruta escura; alguns como a visão atrofiam, outros como os ligados ao paladar expandem-se consideravelmente. Numa tradução livre de um pequeno excerto da introdução do artigo de Tamara Franz-Odendaal:Na biologia evolucionária uma forma de pensar acerca da modularidade é considerar que o organismo em adaptação como sendo constituído por um número de unidades funcionais morfológicas ou morfogenéticas. Por exemplo, um organismo pode ser encarado como tendo módulos dos membros, módulos dos olhos, módulos dos ouvidos, etc.
E um pouco mais adiante:Os peixes cegos das grutas fornecem um exemplo interessante pois indivíduos nalgumas populações perderam a visão e ao mesmo tempo expandiram dois outros orgãos sensoriais: as papilas gustativas e sistema mecanossensitivo da linha lateral.
O sistema de linha lateral é um orgão dos peixes, que consiste em linhas de células ao longo do corpo dos peixes, das guelras até à base da cauda, e que permite ao peixe sentir pequenas diferenças de pressão na água circundante. Este orgão permite ao peixe uma forma de "ecolocação lateral" que lhe permite evitar obstáculos, detectar presas ou evitar potenciais predadores. Os receptores que permitem aos peixes este sentido, designados neuromastros, são grupos de pêlos envolvidos por uma cúpula gelatinosa, com comprimentos da ordem dos décimos de mm. Os peixes cegos das grutas apresentam milhares de neuromastros superficiais, cobrindo quase todo o corpo, enquanto nas formas de superfície estas estruturas são muito mais raras. Para além disso, as cúpulas desses neuromastros têm praticamente o dobro do comprimento das dos peixes da superfície e a estrutura interna dos neuromastros é também diferente, tendo sido modificada de formas que podem aumentar a sensibilidade dos neuromastros. Os módulos gustativos também são muito mais abundantes nos peixes das grutas, em espécial na superfície ventral do crânio, e a importância deste sentido para os peixes das grutas pode ser avaliada notando que a parte do cérebro que processa o cheiro e paladar encontra-se aumentada em cerca de 40% nas formas cegas das grutas quando comparadas com as formas de superfície.
O artigo de Tamara Franz-Odendaal e Brian Hall parte então para uma discussão mais aprofundada dos mecanismos genéticos subjacentes, que eu não vou reproduzir aqui, pois faz referência a um grande volume de literatura especializada, mas o que é importante notar é que o Astyanax mexicanus não é simplesmente um peixe degenerado que pode viver sem visão porque habita num local onde os olhos são inúteis. Pelo contrário, as formas das grutas mostram adaptações notáveis ao seu ambiente, e se alguns sentidos se apresentam atrofiados outros expandiram-se consideravelmente. Na verdade um trabalho mais antigo, de Theresa Burt de Perera (ref2), na revista Proceedings of the Royal Society of London B, mostra outro aspecto notável da sardina de Mexico. As formas cegas mantêm um mapa do ambiente circundante no seu cérebro. Numa tradução livre do resumoOs animais têm muitas vezes que se orientar através de áreas muito maiores que o seu alcance perceptual. O peixe mexicano das cavernas depende da detecção de pequenas perturbações ondulatórias nos objectos através do uso do seu orgão de linha lateral. O seu alcance perceptual (igual ou inferior a 0.05 m) é excedido grandemente pelas suas exigências ecológicas (por volta de 30m). Embora se saiba que possui um mapa espacial do seu ambiente, não se sabe como este peixe liga os diferentes locais (ou a área coberta pelo alcance perceptual).
A questão tal como está posta no resumo é simples. O alcance da linha lateral é de apenas alguns cm, o peixe não consegue ter informação para lá dessa distância. Ora o peixe precisa de explorar um espaço com um raio de cerca de 30 m para sobreviver. A questão é se o peixe sabe onde está, isto é, será que o peixe sabe que aquele espaço de alguns cm onde se encontra fica em tal ou tal lugar na gruta?Usando a propensão dos peixes das grutas para acelerarem quando face a objectos ou ambientes que reconhecem como novos, utilizei um teste comportamental para verificar se os peixes podem aprender e decorar a ordem de elementos numa paisagem.
Quando colocados num ambiente que conhecem bem os peixes deslocam-se calma e lentamente. Quando confrontados quando com paisagens desconhecidas os peixes mudam de comportamento e passam a deslocar-se depressa, o que parece estar relacionado com a forma como constroem os seus mapas cerebrais. A questão é se os peixes constroem um mapa global em que a sequência de estruturas na paisagem é importante, ou se indentificam apenas os locais isoladamente. Ou seja, se os peixes sabem se um local embora familiar não devia estar ali. O que a autora fez foi mudar a ordem de alguns objectos na paisagem, para ver se os peixes reagiam mudando a velocidade a que se deslocavam. A resposta foi sim, os peixes são sensíveis não só ao padrão mas também à ordem em que as estruturas aparecem.Mostro, pelo que sei pela primeira vez, que os peixes cegos mexicanos das grutas podem codificar ordem nos seus mapas espaciais. A habilidade para representar a ordem na qual uma série de locais estão espacialmente ligados é uma ferramenta poderosa para animais que se devem orientar para lá do limite do seu alcance perceptual. O mapa espacial resultante é análogo a um puzzle quebracabeças, onde cada peça representa um local cujo tamanho é constrangido pelo alcance perceptual do animal.
Os seja, cada vez que se deslocam estes peixes estão a resolver um quebra-cabeças com dimensões de dezenas de m, tentando encaixar peças de alguns cm. Trata-se de um feito notável para uma criatura tão pequena. O Astyanax mexicanus das grutas é de facto um organismo magistralmente adaptado ao seu meio ambiente.
Referências
(ref1) Franz-Odendaal, Tamara A. & Hall, Brian K. (2006). Modularity and sense organs in the blind cavefish, Astyanax mexicanus. Evolution & Development 8 (1), 94-100. doi: 10.1111/j.1525-142X.2006.05078.x
(ref2) Burt de Perera, T. (2004). Fish can encode order in their spatial map. Proc. Roy. Soc. Lond. B. 271, 2131-2134.
segunda-feira, fevereiro 13, 2006
Regresso aos mamutes

 O ano de 2006 parece ser de facto o ano do mamute. Para além dos dois artigos citados na semana passada, um outro artigo recente focou também o genoma do mamute. Mas este artigo não se debruçou sobre o ADN mitocondrial mas sim sobre o material genético do núcleo da célula, isto é, os genes que de facto determinam as características físicas do animal. [... ler mais]
O ano de 2006 parece ser de facto o ano do mamute. Para além dos dois artigos citados na semana passada, um outro artigo recente focou também o genoma do mamute. Mas este artigo não se debruçou sobre o ADN mitocondrial mas sim sobre o material genético do núcleo da célula, isto é, os genes que de facto determinam as características físicas do animal. [... ler mais]
Um artigo de Hendrik Poinar e colegas na revista Science de Janeiro (ref1), apresenta este trabalho revolucionário. Numa tradução livre do resumo:Sequenciámos 28 milhões de pares de bases de ADN numa abordagem metagenómica usando uma amostra de mamute lanudo (Mammuthus primigenius) obtida na Siberia. Graças à preservação excepcional da amostra e à utilização de uma nova cadeia de reação de emulsão de polimerase e técnicas de pirossequenciação, identificámos como sendo ADN de mamute 13 milhões de pares de bases (45.4%).
Os termos técnicos algo complicados descrevem apenas o nome dado a este tipo de técnicas que se destinam a reconstruir o genoma a partir de pequenas amostras fragmentadas, e contaminadas por ADN de organismos do solo, e dos próprios investigadores. O importante é a quantidade gigantesca de informação que se conseguiu extrair. No ADN mitocondrial investigado pelos autores dos outros artigos tratava-se de identificar apenas 16,000 pares de base. No núcleo duma célula a quantidade de informação é muito maior: com estes 13 milhões de pares de bases os autores do artigo conseguiram reconstituir apenas cerca de 1% do genoma nuclear. Mas este material genético é importante, pois é ele que nos diz o que faz de um mamute um mamute. Ao compararem o ADN do mamute com o de um parente próximo, o elefante africano, os autores verificaram que estes animais estão de facto muito próximos, tendo tido um antepassado comum há uns poucos milhões de anos:A semelhança entre os nossos dados e o elefante africano (Loxodonta africana) foi 98.55%, consistente com a data de divergência baseada na paleontologia, de 5 a 6 milhões de anos.
 Os autores não fizeram comparações com o elefante asiático, que nos outros estudos mostrou ser o parente mais próximo do mamute, mas não se esperam surpresas na relação entre as três espécies. Aliás, quando se repara no pequeno e felpudo exemplar asiático mostrado aqui ao lado, não é difícil perceber quão próximo estes seres estarão dos seus primos lanudos. E quem sabe, talvez possam num futuro mais ou menos próximo voltar a partilhar a Terra com eles. As últimas linhas do resumo do artigo de Poinar e colegas dizem:
Os autores não fizeram comparações com o elefante asiático, que nos outros estudos mostrou ser o parente mais próximo do mamute, mas não se esperam surpresas na relação entre as três espécies. Aliás, quando se repara no pequeno e felpudo exemplar asiático mostrado aqui ao lado, não é difícil perceber quão próximo estes seres estarão dos seus primos lanudos. E quem sabe, talvez possam num futuro mais ou menos próximo voltar a partilhar a Terra com eles. As últimas linhas do resumo do artigo de Poinar e colegas dizem:A amostra inclui uma quantidade surpeendentemente pequena de ADN ambiental. A alta percentagem de ADN endógeno recuperada deste único mamute deveria permitir a obtenção do seu genoma completo, iniciando o domínio da paleogenómica.
Poinar e colegas estimam que será preciso cerca de um ano até ter o genoma completo do mamute lanudo. Mas isso não significa que se deva pensar em trazer de volta o mamute dentro de pouco tempo. Para já o essencial é assegurar a sobrevivência das espécies que ainda temos, mas quem sabe, talvez no futuro possamos trazer de volta algumas da megafauna extinta.
Para terminar não resisto a mostrar mais uma imagem do pequenino e irrequieto Kandula. Para mais fotos de elefantes, e não só, da autoria de Jessie Cohen, basta consultar as páginas do Smithsonian National Zoological Park aqui.
Referências:
(ref1)Poinar et al. (2006). Metagenomics to Paleogenomics: Large-Scale Sequencing of Mammoth DNA. Science 20, 392-394, Laço DOI.
![]()
domingo, fevereiro 12, 2006
Uma história de luz e de trevas

 O Astyanax mexicanus, conhecido também como "sardina de Mexico" é um pequeno peixe de água-doce, com um comprimento máximo de 12cm, que vive nalguns rios do México e do Sul dos Estados Unidos da América. As populações deste peixe que vivem à superfície possuem olhos grandes, enquanto as populações de um grande número de grutas são cegas e com vários graus de despigmentação (albinas). A sardina de Mexico é um dos poucos exemplos em que o estado funcional de um orgão (nos peixes de superfície) e o estado vestigial (nos peixes cegos) podem ser estudados na mesma espécie. Uma característica da sardina de Mexico que a torna particularmente adaptada a este tipo de estudos, é que se trata de um peixe que se adapta bastante bem ao cativeiro, sendo mesmo razoavelmente popular entre os entusiastas dos aquários.[... ler mais]
O Astyanax mexicanus, conhecido também como "sardina de Mexico" é um pequeno peixe de água-doce, com um comprimento máximo de 12cm, que vive nalguns rios do México e do Sul dos Estados Unidos da América. As populações deste peixe que vivem à superfície possuem olhos grandes, enquanto as populações de um grande número de grutas são cegas e com vários graus de despigmentação (albinas). A sardina de Mexico é um dos poucos exemplos em que o estado funcional de um orgão (nos peixes de superfície) e o estado vestigial (nos peixes cegos) podem ser estudados na mesma espécie. Uma característica da sardina de Mexico que a torna particularmente adaptada a este tipo de estudos, é que se trata de um peixe que se adapta bastante bem ao cativeiro, sendo mesmo razoavelmente popular entre os entusiastas dos aquários.[... ler mais]
O desenvolvimento dos olhos inicia-se da mesma forma nos embriões do peixes das grutas e dos peixes da superfície, mas nas formas cegas as células da lente do olho sofrem morte celular programada, e o olho acaba por ser coberto por uma camada de pele. Experiências em que se transplantou a lente de um peixe da superfície para uma forma cega mostraram que se conseguia induzir desse modo o desenvolvimento do olho (ref1). O Astyanax tem sido utilizado para estudar os mecanismos de morte celular na lente dos olhos e o papel que certos genes desempenham no processo de degenerescência dos olhos nos peixes das grutas. O facto de se dispor de um número variado de populações que provavelmente perderam a visão de forma independente permite ainda ver se os peixes seguiram os mesmos processos de forma paralela ou se diferentes populações seguiram caminhos diferentes. Dois estudos sobre esta simpática criatura acabam de ser publicados na revista Evolution & Development.
Luis Espinasa e William Jeffery (ref2) mostraram que as formas cegas da sardina de Mexico mantêm movimentos retinomotores em resposta às variações de luminosidade idênticos aos peixes de superfície. Para compreender melhor a importância do estudo, nada melhor do que traduzir um dos parágrafos da introdução do artigo:A perda ou redução de características ancestrais é um acontecimento habitual na história evolutiva dos organismos. Exemplos clássicos incluem a regressão dos membros inferiores nas baleias, dentes nas aves, e olhos e pigmentação de peixes em grutas. Dado tempo suficiente, a decadência e perda definitiva de uma característica morfológica é esperada quando se dá a relaxação da seleção natural nesse traço. Contudo, quando um orgão regride durante a evolução, é muitas vezes pouco claro se a forma estrutural ou a função fisiológica se perde primeiro, embora os estudo de caracteres degenerados em animais de grutas geralmente assumam que a característica perdida não é funcional. Para além disso, não se compreende se a perda de função fisiológica é um prerrequisito para a degenerescência funcional de um orgão.
A função que Espinasa e Jeffery escolheram testar foi a presença/ausência de resposta retinomotora nos peixes das grutas. As células epiteliais da retina seguem um padrão de variação de pigmentação que é alterado em função do cliclo diário de variação da luminosidade entre entre o dia e a noite. Embora nos peixes das grutas a retina seja apenas vestigial, e as células epiteliais tenham perdido a pigmentação, é ainda possível seguir alterações de funcionamento. No fundo a questão é se o peixe perderá primeiro toda a funcionalidade associada ao olho, incluindo a presença de um padrão retinomotor, e só depois o olho atrofia, ou se o orgão atrofiou sem que fosse necessário essa perda de funcionalidade. O procedimento adoptado foi o seguinte:Cardumes de embriões de uma só ninhada de peixes de superfície e das grutas foram criados por 2,5,6, e 70 semanas e sujeiitos a um ciclo de 14:10 luz/escuridão (LE) [...] Os animais foram sacrificados a cada 12 horas, começando no meio do intervalo de luz do ciclo LE, fixando-os em 4% de paraformaldeído.
Ou seja, os animais das grutas foram sujeitos a níveis de iluminação ao mesmo tempo e nas mesmas circunstâncias dos peixes da superfície. Os autores testaram ainda variações na iluminação para ver como os peixes respondiam a padrões de luz/escuridão variáveis:cardumes foram criados até ao estádio de 5 semanas sob um ciclo LE 14:10 e então separados em três grupos. O primeiro grupo foi amntido em escuridão constante (EE), o segundo grupo foi mantido em luz constante (LL), e o terceiro grupo foi transferido para um ciclo LE 12:12 que tinha sido atrasado por 12 h. Os animais foram fixados com 4% PFA para histologia a cada 12h, tal como descrito acima,
A análise dos animais da superfície e das grutas mostrou que ambos os grupos seguiam os mesmos tipos de actividade retinal, ou seja ambos tinham o mesmo tipo de resposta retinomotora, quer antes, quer após o início da degenerescência dos olhos nos peixes das grutas. Os autores concluem então queAparentemente, os sinais que regulam a degenerescência do olho são independentes e não entram em conflito com o controlo dos movimentos retinomotores.
Ou seja, durante o processo evolutivo, os peixes das grutas não ficaram necessariamente "funcionalmente cegos" antes da degenerescência do olhos se ter iniciado. Mas o pequeno peixe tem mais segredos que serão focados depois de amanhã, noutra contribuição.
Referências
(ref1) Yamamoto, Y., and W. R. Jeffery. (2002). Probing vertebrate eye development by lens transplantation. Methods 28: 420-426.
(ref2) Espinasa, Luis & Jeffery, William R (2006). Conservation of retinal circadian rhythms during cavefish eye degeneration. Evolution & Development 8 (1), 16-22. doi: 10.1111/j.1525-142X.2006.05071.x
sábado, fevereiro 11, 2006
Mais sobre mamutes lanudos e elefantes

 Pelos vistos os mamutes estão realmente na moda. A Nature de Fevereiro traz também um artigo sobre a determinação do ADN mitocondrial do mamute lanudo, usando uma técnica que permite utilizar uma quantidade infíma de material orgânico. Esse é um progresso importante pois contrariamente à percepção popular os mamutes que se encontram congelados estão em geral em péssimas condições. [... ler mais]
Pelos vistos os mamutes estão realmente na moda. A Nature de Fevereiro traz também um artigo sobre a determinação do ADN mitocondrial do mamute lanudo, usando uma técnica que permite utilizar uma quantidade infíma de material orgânico. Esse é um progresso importante pois contrariamente à percepção popular os mamutes que se encontram congelados estão em geral em péssimas condições. [... ler mais]
O novo método para reconstituir o código genético é descrito num artigo de Johannes Krause e colegas (ref1). Numa tradução livre do resumo:Ao estudar os genomas de espécies extintas, as duas limitações principais são a pequena quantidade de ADN antigo endógeno, e o seu estado de degradação, embora produtos de até 1,600 pares de bases (pb) tenham sido amplificados nalguns casos raros.
Os pares de bases são as unidades que compõem o ADN. No caso dos mamutes e elefantes o genoma das mitocôndrias corresponde a qualquer coisa como 16,000 pares de bases. O problema é que em amostras de grande antiguidade o genoma está partido em muitos bocados mais pequenos. Continuando:Utilizando reações de polimerase em cadeia com algum pequenas sobreposições, sequências maiores ou mesmo genomas mitocondriais completos podem ser reconstruídos, mas esta aproximação é limitada pelo pequeno número de amplificações que se podem fazer a partir de amostras escassas. Logo, mesmo em espécies do Pleistoceno bem estudadas como mamutes, preguiças terrícolas e ursos das cavernas, não se recuperaram sequências com mais de 1000 pb. Relatamos aqui a sequência completa do genoma mitocondrial do mamute do Pleistoceno Mammuthus primigenius. Utilizámos cerca de 200 mg de osso e uma abordagem nova que permite a recuperação simultânea de várias sequências a partir de pequenas amostras de ADN degradado.
Ou seja, usando alguns dos bocados de ADN e acertando as "pontas" com outros fragmentos consegue-se por vezes reconstruir sequências bastante grandes. No fundo é como se tivéssemos muitas cópias de um texto, as cortássemos em bocados de tamanhos aleatórios, e depois procurássemos fragmentos com sequência de palavras semelhantes. Se tivermos um grande número de cópias há uma grande probabilidade de os fragmentos "encaixarem" e conseguirmos reconstituir todo texto. O problema com as amostras de ADN muito antigas é que em geral a quantidade de material que ainda é aproveitável (não degradado) é relativamente pequena. O que os investigadores conseguiram foi aumentar a eficácia do processo, e assim a partir de uma quantidade realmente diminuta de material orgânico foi possível reconstruir uma sequência completa. Para finalizar:A nossa análise da filogenia mostra que o mamute estava mais estreitamente relacionado com o elefante asiático que com o elefante africano. Contudo, a divergência entre o mamute e os elefantes asiático e africano ocorreu num período de tempo bastante curto, correspondendo a apenas cerca de 7% da extensão da árvore filogenética para as três linhagens.
Os resultados que obtêm são semelhantes aos obtidos pelos autores do estudo de que se falou ontem. Se pudéssemos retroceder no tempo, existiria um período em que os antepassados do mamute e elefantes actuais formariam uma única espécie. Se em seguida avançássemos um pouco a partir desse momento veríamos que esses indivíduos teriam dado origem a duas espécies, uma que viria a dar os elefantes africanos actuais, e uma outra que se separaria pouco tempo depois em antepassados apenas dos mamutes e antepassados apenas dos elefantes asiáticos actuais. O estudo da Nature mostrou que o processo de especiação foi no entanto muito rápido, umas poucas centenas de milhares de anos apenas.
Para terminar, e já que o elefante asiático foi amplamente publicitado na última contribuição, eis aqui uma imagem da simpática variante africana.
Referências
(ref1) Multiplex amplification of the mammoth mitochondrial genome and the evolution of Elephantidae. Johannes Krause, Paul H. Dear, Joshua L. Pollack, Montgomery Slatkin, Helen Spriggs, Ian Barnes, Adrian M. Lister, Ingo Ebersberger, Svante Pääbo and Michael Hofreiter. Laço DOI.
sexta-feira, fevereiro 10, 2006
Primos não muito afastados

 Não, nenhum elefante foi magoado ao tirar estas fotografias, e a imagem não é tão macabra como parece. O elefante asiático, Elephas maximus, está apenas a limpar o olho e não a chorar a perda de um dos seus membros. Aliás, a perna da esquerda nem pertence a um elefante asiático. Trata-se da reconstrução da perna de um Mammuthus primigenius, mais conhecido por mamute lanudo, e foi baseada em achados com 33,750 anos. [... ler mais]
Não, nenhum elefante foi magoado ao tirar estas fotografias, e a imagem não é tão macabra como parece. O elefante asiático, Elephas maximus, está apenas a limpar o olho e não a chorar a perda de um dos seus membros. Aliás, a perna da esquerda nem pertence a um elefante asiático. Trata-se da reconstrução da perna de um Mammuthus primigenius, mais conhecido por mamute lanudo, e foi baseada em achados com 33,750 anos. [... ler mais] O achado original que se mostra aqui à esquerda tinha na verdade um aspecto muito mais ressequido. As condições de conservação desta perna, encontrada no nordeste da Sibéria em 1986, eram contudo suficientemente boas para que ADN mitocondrial pudesse ser extraído dos músculos e não apenas do osso como é mais habitual. Foi isso que fizeram Rogaev e colegas, que publicaram os resultados no PLoS Biology (ref1). Numa tradução livre do resumo:
O achado original que se mostra aqui à esquerda tinha na verdade um aspecto muito mais ressequido. As condições de conservação desta perna, encontrada no nordeste da Sibéria em 1986, eram contudo suficientemente boas para que ADN mitocondrial pudesse ser extraído dos músculos e não apenas do osso como é mais habitual. Foi isso que fizeram Rogaev e colegas, que publicaram os resultados no PLoS Biology (ref1). Numa tradução livre do resumo:As relações filogenéticas entre o mamute lanudo (Mammuthus primigenius), e os elefantes asiático (Elephas maximus) e africano (Loxodonta africana) permanecem por esclarecer. Relatamos aqui a sequência completa do genoma mitrocondrial (16,842 pares de bases) de um mamute lanudo extraído do restos preservados em permafrost da época do Pleistoceno, a sequência mitrocondrial mais antiga determinada até à data.
Os últimos meses têm sido ricos em estudos genéticos de restos de mamutes, mas esses estudos têm recorrido a fragmentos relativamente menores, este estudo baseia as suas conclusões no genoma completo das mitocôndrias. As mitocôndrias são pequenas estruturas que existem no interior das nossas células (bem como dos outros animais e plantas) que têm um papel fulcral na respiração. São herdadas apenas da mãe e possuem o seu próprio código genético relativamente simples mas que é suficiente para determinar relações de parentesco na linha materna. O facto de possuirem uma sequência completa e de boa qualidade permitiu aos investigadores determinar o parentesco entre o mamute e as espécies de elefante usando amostras de ADN mitocondrial desses animais. Voltando ao resumo:A reconstrução da filogenia do clâ Elephantinae sugere que M. primigenius e E. maximus são espécies irmãs que divergiram um pouco depois do seu antecessor comum se ter separado da linhagem L. africana.
Ou seja, se voltássemos atrás no tempo encontraríamos um período em que os antecessores do mamute e do elefante asiático fariam parte da mesma espécie, mas os antepassados do elefante africano seriam já uma espécie separada. Os mamutes lanudos eram portanto realmente apenas elefantes com abundância de pêlo. Em jeito de despedida mais uma fotografia de um elefante asiático, que esperemos não siga o caminho do seu primo lanudo: a extinção.
Referências
(ref1) Rogaev EI, Moliaka YK, Malyarchuk BA, Kondrashov FA, Derenko MV, et al. (2006). Complete Mitochondrial Genome and Phylogeny of Pleistocene Mammoth Mammuthus primigenius. PLoS Biol 4(3): e73. Laço DOI.
quinta-feira, fevereiro 09, 2006
O pequeno tiranossauro com a crista bizarra
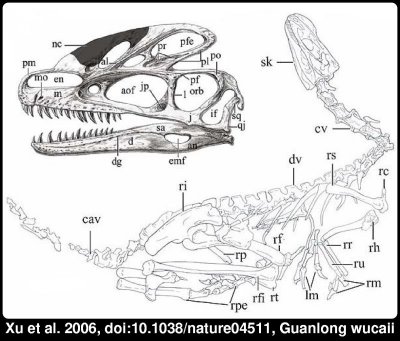
 Curiosamente o espinossauro de que se fala na contribuição abaixo foi ignorado pela imprensa, que publicitou no entanto largamente a descoberta de um parente afastado do tiranossauro, o Guanlong wucaii, com uns míseros 3 metros de comprimento e que viveu cerca de 100 milhões antes do seu primo maior. [... ler mais]
Curiosamente o espinossauro de que se fala na contribuição abaixo foi ignorado pela imprensa, que publicitou no entanto largamente a descoberta de um parente afastado do tiranossauro, o Guanlong wucaii, com uns míseros 3 metros de comprimento e que viveu cerca de 100 milhões antes do seu primo maior. [... ler mais]
Esta criatura é descrita em detalhe num artigo de Xu e colegas(ref1). Trata-se de uma descoberta interessante por vários motivos. Por um lado pela idade do achado, pois vem esclarecer a relação entre as diversas famílias de animais que constituem aquilo que se chama os tiranossaurídeos. Mas o que chama mais a atenção neste dinossauro é a estranha crista no alto da cabeça. Visto que uma estrutura frágil deste tipo parece de pouca utilidade para um predador, os autores do artigo da sugerem que se trataria de uma estrutura usada para atrair membros do sexo oposto, à semelhança da cauda de um pavão. Uma reconstituição artística do aspecto que este animal poderia ter pode ser encontrada aqui.
A reconstituição destes animais com penas longas nas asas e cauda, bem como penas menores por todo o corpo não é completamente especulativa, pois descobertas recentes mostraram que parentes próximos dos tiranossaurídeos possuíam de facto penas. A maioria dos autores considera mesmo que as aves são dinossauros. A minha reconstrução favorita de um dinossauro é DEINONYCHUS: A NEW LOOK FOR THE YEAR 2000 de Luis Rey, que mostra um parente próximo dos famosos velociraptores do Parque Jurássico com um aspecto que faz pensar numa galinha com mau feitio.
Referências
(ref1) Xing Xu, James M. Clark, Catherine A. Forster, Mark A. Norell, Gregory M. Erickson, David A. Eberth, Chengkai Jia and Qi Zhao (2006). A basal tyrannosauroid dinosaur from the Late Jurassic of China. Nature 439, 715-718. Laço DOI.
Chega-te para lá tiranossauro
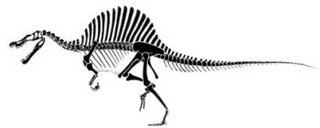
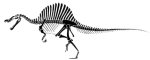 É oficial, o Tirannossaurus rex foi ultrapassado como o maior predador terrestre, e por uma larga margem. Todos aqueles que viram o Parque Jurássico III se lembram que o vilão da história era um estranho dinossauro com uma cabeça que fazia lembrar um crocodilo e com uma espécie de vela nas costas, o Spinosaurus aegyptiacus. [... ler mais]
É oficial, o Tirannossaurus rex foi ultrapassado como o maior predador terrestre, e por uma larga margem. Todos aqueles que viram o Parque Jurássico III se lembram que o vilão da história era um estranho dinossauro com uma cabeça que fazia lembrar um crocodilo e com uma espécie de vela nas costas, o Spinosaurus aegyptiacus. [... ler mais]
Os restos algo incompletos do Spinosaurus aegyptiacus foram descobertos em 1912 pelo coleccionador Markgraf, no vale de Baharija no Egipto, tendo sido descritos por Strommer em 1915. Esses restos acabariam por se perder durante a guerra na Europa, mas novos exemplares foram descobertos nos últimos anos. Exemplares esses que sugeriam que o espinossauro poderia facilmente destronar outros superpredadores do passado da posição de maior carnívoro terrestre. Isso acaba de ser comprovado num artigo de Dal Sasso e colegas, no Journal of Vertebrate Paleontology (ref1). Tradução livre do resumo:Novos exemplares do estranho terópode Spinosaurus cf. S. aegyptiacus do Cretácico tardio (Cenomaniano inicial) de Marrocos revelam nova informação acerca da estrutura do rosto e sobre o grande tamanho atingido pelos adultos da espécie.
O período indicado significa que os espinossauros viveram cerca de 30 milhões de anos antes dos tiranossauros e noutra região do globo, pelo que um combate como o do filme nunca poderia ter acontecido. Um pouco mais adiante:O tamanho do rosto sugere que o Spinosaurus pode muito bem ter excedido o tamanho dos adultos de outros terópodes como o Tirannossaurus e o Giganotosaurus.
O giganotosauro era um predador da Argentina que tinha cerca de 15 metros de comprimento e já superava o tiranossauro em qualquer coisa como um metro e meio. Os autores do artigo descrevem em detalhe um fragmento do crânio do espinossauro. Embora esse fragmento tenha "apenas" um metro de comprimento, a comparação com outros crânios conhecidos indica que o crânio completo teria uma dimensão total de 1.75 metros. Isto é, o crânio desta criatura era do tamanho dum homem adulto. Baseados nas dimensões do crânio e no conhecimento de outros esqueletos desta criatura os cientistas calculam que o indivíduo de ondem provêm estes restos teria de 16 a 18 metros de comprimento, claramente o maior predador que andou pela superfície da Terra e que deixou algum tipo de vestígios.
Referências
(ref1) Dal Sasso, C., Maganuco, S., Buffetaut, E. & Mendezm M. A. 2005. New information on the skull of the enigmatic theropod Spinosaurus, with remarks on its size and affinities. Journal of Vertebrate Paleontology, 25, 888-896.
quarta-feira, fevereiro 08, 2006
Ave-do-paraíso perdida descoberta no paraíso

 Há imagens que practicamente dispensam palavras. Esta ave-do-paraíso foi recentemente redescoberta por uma expedição conduzida por Bruce Beehler, vice-presidente do Conservation International, numa expedição às montanhas Foja, uma região remota na parte ocidental da ilha da Nova-Guiné (Indonésia). [... ler mais]
Há imagens que practicamente dispensam palavras. Esta ave-do-paraíso foi recentemente redescoberta por uma expedição conduzida por Bruce Beehler, vice-presidente do Conservation International, numa expedição às montanhas Foja, uma região remota na parte ocidental da ilha da Nova-Guiné (Indonésia). [... ler mais]
A expedição foi um sucesso, um grande número de novas espécies foram identificadas, incluindo rãs, borboletas e a ave que se mostra abaixo. Este é um mundo onde se encontram cangurus arborícolas, mamíferos com bico e que põem ovos (equidnas), e muitas espécies de aves-do-paraíso, e nenhum destes animais apresenta medo dos seres humanos. Por exemplo, as aves-do-paraíso machos executavam os bailados mesmo em frente aos cientistas. Deve ser realmente um local mágico. O título desta contribuição é emprestado de um texto encontrado no blog Living the Scientific Life (Scientist Interrupted). O texto, escrito por uma ornitóloga actualmente à procura de emprego, está em inglês e inclui uma série de laços relevantes. Recomendo-o vivamente.
O título desta contribuição é emprestado de um texto encontrado no blog Living the Scientific Life (Scientist Interrupted). O texto, escrito por uma ornitóloga actualmente à procura de emprego, está em inglês e inclui uma série de laços relevantes. Recomendo-o vivamente.
terça-feira, fevereiro 07, 2006
Os camarões do antártico

 A criatura de hoje não é tão sinistra como as que apareceram nas contribuições mais recentes. Com o nome científico de Euphausia superba, mais conhecidas pela designação norueguesa de krill, estas pequenas criaturas (cerca de 6cm de comprimento) são a espécie animal com maior sucesso à face da Terra. Estima-se que a biomassa do krill do antártico ande algures entre 125 a 725 milhões de toneladas. Muitas pessoas conhecem o krill apenas porque se trata do alimento preferido de algumas espécies de baleias e focas. Na verdade estes minúsculos parentes dos camarões são muito importantes para o equilíbrio do planeta, e não apenas uma abundante fonte de alimento para animais maiores. [... ler mais]
A criatura de hoje não é tão sinistra como as que apareceram nas contribuições mais recentes. Com o nome científico de Euphausia superba, mais conhecidas pela designação norueguesa de krill, estas pequenas criaturas (cerca de 6cm de comprimento) são a espécie animal com maior sucesso à face da Terra. Estima-se que a biomassa do krill do antártico ande algures entre 125 a 725 milhões de toneladas. Muitas pessoas conhecem o krill apenas porque se trata do alimento preferido de algumas espécies de baleias e focas. Na verdade estes minúsculos parentes dos camarões são muito importantes para o equilíbrio do planeta, e não apenas uma abundante fonte de alimento para animais maiores. [... ler mais] Embora a dieta destas criaturas possa ser algo variada incluindo pequenos seres que fazem parte do zooplâncton, e até outros membros da sua espécie, o krill alimenta-se sobretudo de infímas diatomáceas (20 micrómetros), que filtra através de uma espécie de cesto que forma com as seis patas dianteiras. Para além de pescar as algas na água o krill pode também raspá-las nas camadas de gelo que cobrem os mares do antártico, deslocando-se em zique-zague como um minúsculo cortador de relva. O krill é um comensal com fracas maneiras à mesa, cuspindo frequemente gandes agregados de algas a que os cientistas chamam "bolas de cuspo". Um dos aspectos mais interessantes dos hábitos alimentares do krill é que a eficiência do processo digestivo é relativamente baixa e uma grande quantidade de carbono é retida nas fezes e libertada juntamente com as carapaças das diatomáceas. Quer os pequenos tubos de fezes quer as bolas de cuspos são pesadas e afundam rapidamente, transportando grandes quantidades de carbono que se vão depositar nos sedimentos do fundo do oceano a profundidades de alguns quilómetros, onde ficam sequestrados por períodos da ordem de 1000 anos. O krill tem assim uma importância fundamental no ciclo do carbono terrestre, pois se as algas forem consumidas por outras criaturas marinhas o carbono fica à superfície. Ora muitos dos aspectos do comportamento do krill e deste mecanismo de reciclagem do carbono ainda são mal compreendidos.
Embora a dieta destas criaturas possa ser algo variada incluindo pequenos seres que fazem parte do zooplâncton, e até outros membros da sua espécie, o krill alimenta-se sobretudo de infímas diatomáceas (20 micrómetros), que filtra através de uma espécie de cesto que forma com as seis patas dianteiras. Para além de pescar as algas na água o krill pode também raspá-las nas camadas de gelo que cobrem os mares do antártico, deslocando-se em zique-zague como um minúsculo cortador de relva. O krill é um comensal com fracas maneiras à mesa, cuspindo frequemente gandes agregados de algas a que os cientistas chamam "bolas de cuspo". Um dos aspectos mais interessantes dos hábitos alimentares do krill é que a eficiência do processo digestivo é relativamente baixa e uma grande quantidade de carbono é retida nas fezes e libertada juntamente com as carapaças das diatomáceas. Quer os pequenos tubos de fezes quer as bolas de cuspos são pesadas e afundam rapidamente, transportando grandes quantidades de carbono que se vão depositar nos sedimentos do fundo do oceano a profundidades de alguns quilómetros, onde ficam sequestrados por períodos da ordem de 1000 anos. O krill tem assim uma importância fundamental no ciclo do carbono terrestre, pois se as algas forem consumidas por outras criaturas marinhas o carbono fica à superfície. Ora muitos dos aspectos do comportamento do krill e deste mecanismo de reciclagem do carbono ainda são mal compreendidos.
Um artigo recente (ref1), saído no Current Biology, da autoria de Geraint tarling e Magnus Johnson, mostra um aspecto curioso nos hábitos alimentares do krill. Após se empaturrarem de diatomáceas à superfície os minúsculos camarões deixam-se afundar até grandes profundidades, onde completam a digestão, libertam os seus tubinhos de fezes e voltam à superfície para se alimentarem de novo. Este novo estudo mostra que no processo de vaivém o krill trasporta muito mais carbono do que se pensava, sendo uma espécie muito mais importante para o equilíbrio global do planeta do que se suspeitava.
Ficha técnica
Imagens de muito maior resolução, em que pormenores do corpo da criatura podem ser facilmente vistos, encontram-se disponíveis na internet, cortesia de Uwe Kils.
Referências
(ref1) Geraint A. Tarling, and Magnus L. Johnson (2006). Satiation gives krill that sinking feeling. Current Biology, Volume 16, Issue 3, Pages R83-R84. Laço DOI.
Olduvai George e estranhos crocodilos
 Numa contribuição anterior, falou-se num estranho animal já extinto, o Effigia okeeffeae, uma espécie de "crocodilo avestruz". Pois bem, o artista Carl Buell, mais conhecido por Olduvai George, colocou no seu blog uma reconstituição dessa criatura, bem como do Chimerosuchus, um crocodilo herbívoro (não, não se trata de um engano) contemporâneo dos dinossauros, e de um crocodilo dos nossos dias ameaçado de extinção, o gavial. As reconstituições podem ser encontradas aqui.
Numa contribuição anterior, falou-se num estranho animal já extinto, o Effigia okeeffeae, uma espécie de "crocodilo avestruz". Pois bem, o artista Carl Buell, mais conhecido por Olduvai George, colocou no seu blog uma reconstituição dessa criatura, bem como do Chimerosuchus, um crocodilo herbívoro (não, não se trata de um engano) contemporâneo dos dinossauros, e de um crocodilo dos nossos dias ameaçado de extinção, o gavial. As reconstituições podem ser encontradas aqui.
quinta-feira, fevereiro 02, 2006
O vírus que engorda

 A imagem ao lado mostra um adenovírus, membro de uma família que provoca doenças em humanos e animais, provocando tosse, espirros e outros sintomas duma constipacão. Esta família de vírus é interessante porque experiências em ratos e galinhas mostraram que alguns deles causam aumentos de peso nos animais. Esses resultados levaram alguns investigadores a sugerir que as pessoas podem contrair a obesidade como quem contrai uma constipação. [... ler mais]
A imagem ao lado mostra um adenovírus, membro de uma família que provoca doenças em humanos e animais, provocando tosse, espirros e outros sintomas duma constipacão. Esta família de vírus é interessante porque experiências em ratos e galinhas mostraram que alguns deles causam aumentos de peso nos animais. Esses resultados levaram alguns investigadores a sugerir que as pessoas podem contrair a obesidade como quem contrai uma constipação. [... ler mais]
Embora a sugestão ainda gere alguma controvérsia não deixa de fazer algum sentido pois a obesidade tem alastrado quase como uma epidemia. Leah Wigham e colegas, num estudo publicado no American Journal of Physiology and Regulatory Integrative Comparative Physiology (ref1), mostraram que a infecção com uma variante particular, odenovírus humano 37 (Ad-37) aumenta significativamente a quantidade de gordura acumulada nos tecidos. Numa tradução livre do resumo:O adenovírus humano-36 (Ad-36) aumenta a adiposidade nas galinhas, ratos e primatas não humanos, e está associado com a obesidade em humanos.
Esta é uma referência a um trabalho anterior (ref2), e que será discutido mais adiante. Wigham e colegas investigam outras estirpes do vírus:Este estudo testou se três outros adenovirus humanos aumentam a adeposidade em galinhas e células 3T3-L1 in vitro. Inoculámos galinhas à idade de 3 semanas com o Ad-2, Ad-31 ou Ad-37. O consumo de alimento e o peso foram registados durante 3 semanas 3 meia, tendo as galinhas sido mortas em seguida[...]
Um pouco macabro, embora talvez um destino um pouco mais digno do que acabar como refeição. Um pouco adiante:A gordura nas vísceras e a gordura corporal total foram significativamente mais elevadas no grupo Ad-37 comparado com todos os outros grupos. Os pesos finais foram mais elevados nas galinhas inoculadas com Ad-37 que nas galinhas com Ad-2, mas não significativamente mais elevados que nas galinhas do grupo de controlo ou no grupo Ad-31. O consumo total de alimento foi o mesmo nos dois grupos.
Em resumo, embora não aumente substancialmente o peso o adenovirus aumenta significativamente a quantidade de gordura nos tecidos das aves. É óbvio que este tipo de estudos não se pode fazer com humanos por razões éticas, mas há formas indirectas de testar a relação entre os adenovírus e a obesidade humana. Isso foi feito num estudo anterior, no artigo indicado como (ref2), para um outro membro da família, o Ad-36. Nesse estudo foi analisado um grupo de 500 voluntários magros e obesos, incluindo um certo número de gémeos. O estudo verificou que cerca de 30% dos indivíduos obsesos mostravam sinais do vírus, contra apenas 10% dos indivíduos não obesos (e apenas 5% dos indivíduos classificados como magros). Embora alvo de debate e alguma controvérsia, estes estudos apontam para uma forma algo desconcertante de combater a obesidade, uma simples vacina. Por outro lado reforçam a aceitação da obesidade como uma verdadeira doença e não apenas uma consequência da mudança de hábitos alimentares e de estilo de vida.
Referências
(ref1) APPETITE, OBESITY, DIGESTION, AND METABOLISM:
Leah D. Whigham, Barbara A. Israel, and Richard L. Atkinson
Adipogenic potential of multiple human adenoviruses in vivo and in vitro in animals. Am J Physiol Regulatory Integrative Comp Physiol, Jan 2006; 290: R190 - R194.
(ref2) Human adenovirus-36 is associated with increased body weight and paradoxical reduction of serum lipids. R L Atkinson, N V Dhurandhar, D B Allison, R L Bowen, B A Israel, J B Albu and A S Augustus. International Journal of Obesity (2005) 29, 281.


