




 Esta fotografia, tirada no Jardim Zoológico de Los Angeles, mostra um chimpanzé, de seu nome científico Pan troglodytes, aparentemente perdido em cogitações. Este chimpanzé pensador tem tudo a ver com a contribuição que escolhi para hoje. Como os três ou quatro leitores habituais deste blog sabem, tenho dedicado alguns dos textos mais recentes à problemática da inteligência animal. Em particular tenho discutido aqui trabalhos sobre a forma como diferentes grupos de animais se comportam nos chamados estudos do deslocamento invisível. [... ler mais]
Esta fotografia, tirada no Jardim Zoológico de Los Angeles, mostra um chimpanzé, de seu nome científico Pan troglodytes, aparentemente perdido em cogitações. Este chimpanzé pensador tem tudo a ver com a contribuição que escolhi para hoje. Como os três ou quatro leitores habituais deste blog sabem, tenho dedicado alguns dos textos mais recentes à problemática da inteligência animal. Em particular tenho discutido aqui trabalhos sobre a forma como diferentes grupos de animais se comportam nos chamados estudos do deslocamento invisível. [... ler mais]
Os chimpanzés também tinham mostrado um nível elevado de sucesso nos testes de deslocamento invisível, e Emma Collier-Baker e colegas decidiram verificar se isso se devia a uma real compreensão do fenómeno ou se, como no caso dos cães, se tratava de uma qualquer estratégia associativa simples. Os resultados desse estudo foram publicados na revista Animal Cognition (ref1) e parecem não deixar grandes dúvidas de que os chimpanzés de facto compreendem este tipo de tarefas, o que colocaria as suas capacidades cognitivas representacionais ao nível das crianças humanas com dois anos de idade.
Eis aqui uma fotografia do dispositivo experimental utilizado no estudo: A bolinha que o investigador tem na mão é o objecto que o chimpanzé deve procurar. Consiste numa guloseima (fruto seco ou alteia) e serve portanto também de recompensa. Já agora, um à-parte. As "crianças" em Portugal desconhecem em geral o que é a alteia e, embora familiarizadas com o termo inglês marshmallow, ficam em geral muito surpeendidas quando lhes digo que aquele doce esponjoso é mesmo aquilo que os americanos assam nas fogueiras. Os chimpanzés pelos vistos gostam.
A bolinha que o investigador tem na mão é o objecto que o chimpanzé deve procurar. Consiste numa guloseima (fruto seco ou alteia) e serve portanto também de recompensa. Já agora, um à-parte. As "crianças" em Portugal desconhecem em geral o que é a alteia e, embora familiarizadas com o termo inglês marshmallow, ficam em geral muito surpeendidas quando lhes digo que aquele doce esponjoso é mesmo aquilo que os americanos assam nas fogueiras. Os chimpanzés pelos vistos gostam.
O pequeno copo abaixo e à esquerda na mesa é onde a guloseima vai ser escondida para o deslocamento. Uma das três caixas é o destino final da bolinha. A posição inicial do dispositivo de deslocamento era escolhida de forma aleatória entre quatro possibilidades: ou entre duas caixas, ou num qualquer dos extremos. O teste padrão é simples: o dispositivo de transporte é usado para levar o objecto atrás da caixa alvo, a tampa atrás da caixa é levantada, e o objecto tansferido lá para dentro sem que o chimpanzé o veja. O dispositivo de transporte é então trazido para a frente da caixa, rodado e levantado ao nível dos olhos do chimpanzé que vê que o objecto não está lá dentro. O dispositivo é depois levado até à sua posição inicial com a abertura para baixo. O investigador pede em seguida ao chimpanzé que indique a posição da guloseima. Se acertar pode ficar com ela. Se falhar mostram-lhe onde estava mas não lhe dão a guloseima. Os autores do estudo fizeram este teste com 4 variantes. Numa tradução livre do resumo:Estudos prévios sugerem que os chimpanzés compreendem o deslocamento invisível único. Contudo, esta tarefa piagetiana pode ser resolvida pela utilização de estratégias de busca simples e não pela representação da trajectória passada de um objecto. Administrámos por isso quatro condições de controlo a dois chimpanzéns por forma a separar estratégias de busca associativas do desempenho devido à representação mental. Estratégias envolvendo a utilização de pistas dadas pelo investigador, procura na última ou primeira caixa visitada pelo dispositivo de deslocamento, e busca nas caixas adjacentes ao dispositivo de deslocamento foram controladas de forma sistemática.
Nalguns testes controlou-se utilização de pistas dadas pelo investigador colocando uma máscara no investigador por forma a que o chimpanzé não conseguisse ver-lhe o rosto. Em seguida seguia-se o procedimento normal. O teste da última caixa era um pouco mais complicado. Seguia-se o procedimento normal, escondendo o objecto e mostrando o copo vazio ao chimpanzé, mas em seguida o copo era levado atrás de outra caixa, simulava-se o movimento de transferência, mostrava-se de novo o copo vazio e colocava-o na posição inicial virado para baixo. O teste da primeira caixa era o inverso. O copo era levado atrás de uma caixa, simulava-se a transferência, mostrava-se o copo ao chimpanzé que via que o objecto ainda estavá lá e fazia-se então a transferência para uma outra caixa, mostrava-se o copo vazio, que era depois colocado com a abertura para baixo na posição inicial. Os controles são importantes pois verificou-se que alguns animais, que pareciam compreender o deslocamento invisível, seguiam estratégias muito simples mas eficazes nalgumas situações. Os tamarins, por exemplo, escolhem na maioria das vezes a última caixa visitada pelo dispositivo de deslocamento, enquanto os cães, por sua vez, escolhem preferencialmente a caixa adjacente ao dispositivo de deslocamento.Os chimpanzés não mostraram indicações de utilizarem essas estratégias simples, sugerindo que as suas capacidades para representarem mentalmente o deslocamento invisível único é comparável ao de crianças de 18 a 24 meses de idade.
O desempenho dos chimpanzés foi quase perfeito. Eis aqui os resultados para os dois chimpanzés para 5 tentativas em quatro versões da tarefa, com a ressalva do dispositivo de deslocamento ser ou não deixado junto à caixa onde o objecto ficou escondido.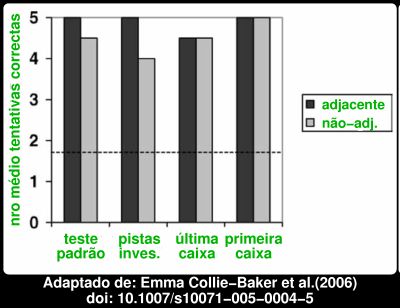 A linha a tracejado representa o que se esperaria se os chimpanzés escolhessem ao acaso. Aliás, um dos chimpanzés, Ockie, mostrou ser particularmente dotado. Eis aqui os resultados por chimpanzé, somando as tentativas com dispositivo adjacente e não adjacente:
A linha a tracejado representa o que se esperaria se os chimpanzés escolhessem ao acaso. Aliás, um dos chimpanzés, Ockie, mostrou ser particularmente dotado. Eis aqui os resultados por chimpanzé, somando as tentativas com dispositivo adjacente e não adjacente: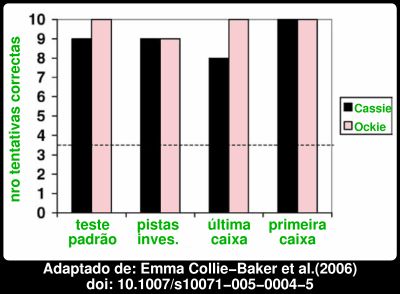 Apenas falhou uma vez em 40 tentativas! Mesmo o Cassie portou-se bastante bem, acertando 90% das vezes, sempre muito acima da linha a tracejado que marca o que se esperaria para uma escolha ao acaso. Já agora, embora o nome Cassie possa induzir em erro, em particular devido à minha escolha do cor-de-rosa para o gráfico, tratava-se de dois machos com 27 e 31 anos. As diferenças com os cães são mais que evidentes no gráfico abaixo, que compara o desempenho de chimpanzés, cães, e crianças, no teste padrão, nos casos em que o dispositivo é deixado adjacente, ou não adjacente à caixa alvo.
Apenas falhou uma vez em 40 tentativas! Mesmo o Cassie portou-se bastante bem, acertando 90% das vezes, sempre muito acima da linha a tracejado que marca o que se esperaria para uma escolha ao acaso. Já agora, embora o nome Cassie possa induzir em erro, em particular devido à minha escolha do cor-de-rosa para o gráfico, tratava-se de dois machos com 27 e 31 anos. As diferenças com os cães são mais que evidentes no gráfico abaixo, que compara o desempenho de chimpanzés, cães, e crianças, no teste padrão, nos casos em que o dispositivo é deixado adjacente, ou não adjacente à caixa alvo. A "batota" dos cães é óbvia. Os outros grandes símios, gorilas e orangotangos também mostraram não utilizar regras associativas simples, e logo também parecem compreender o conceito. Alguns autores sugerem por isso que a capacidade deveria existir no antepassado comum dos grandes antropóides e do homem, há mais de 14 milhões de anos atrás.
A "batota" dos cães é óbvia. Os outros grandes símios, gorilas e orangotangos também mostraram não utilizar regras associativas simples, e logo também parecem compreender o conceito. Alguns autores sugerem por isso que a capacidade deveria existir no antepassado comum dos grandes antropóides e do homem, há mais de 14 milhões de anos atrás.
Embora os chimpanzés consigam um bom desempenho nestas tarefas de deslocamento invisível e revelem capacidade mentais de representação secundária dos objectos, falham quando se fazem testes de deslocamento invisível duplo. Nestes testes o dispositivo de deslocamento visita duas caixas não adjacentes, e falarei deles dentro de alguns dias.
Ficha técnica
Imagem do chimpanzé pensativo no início da contribuição cortesia de Aaron Logan, retirada da sua galeria LIGHTmatter.
Referências
(ref1) Emma Collie-Baker, Joanne M. Davis, Mark Nielsen and Thomas Suddendorf (2006). Do chimpanzees (Pan troglodytes) understand single invisible displacement? Animal Cognition, Volume 9, 55-61. Laço DOI.
quarta-feira, agosto 30, 2006
Como achar guloseimas que se deslocam invisíveis
terça-feira, agosto 29, 2006
A cheirar o caminho de volta

 Esta imagem mostra "borrachos", nome vulgar dos jovens (com menos de 4 semanas) de uma criatura cujo nome científico é Columba livia. Neste caso trata-se de recém-nascidos no seu primeiro dia de vida. Creio que toda a gente identificou o ser em questão, o pombo, aquele animal que evoluiu com a missão expressa de emporcalhar as estátuas nos parques das nossas cidades. Embora não tenha grande estima pelos adultos, os pequenos sempre me fascinaram. São tão feios que acabam por ser irresistíveis. Vou hoje falar de um dos aspectos mais notáveis dessas aves, a facilidade que mostram em voltar a casa mesmo quando largados a grandes distâncias. Esta habilidade dos pombos correios nunca foi convenientemente explicada, embora os cientistas tenham avançado com algumas hipóteses. Por exemplo, sabia-se que os pombos são sensíveis aos campos magnéticos, e suspeitava-se que os pombos se guiavam pelo campo magnético terrestre. Ora o facto de se poder fazer algo nem sempre quer dizer que se faça, e pelos vistos os pombos usam um sentido menos misterioso, e mais familiar para nós, para encontrarem o caminho de casa. [... ler mais]
Esta imagem mostra "borrachos", nome vulgar dos jovens (com menos de 4 semanas) de uma criatura cujo nome científico é Columba livia. Neste caso trata-se de recém-nascidos no seu primeiro dia de vida. Creio que toda a gente identificou o ser em questão, o pombo, aquele animal que evoluiu com a missão expressa de emporcalhar as estátuas nos parques das nossas cidades. Embora não tenha grande estima pelos adultos, os pequenos sempre me fascinaram. São tão feios que acabam por ser irresistíveis. Vou hoje falar de um dos aspectos mais notáveis dessas aves, a facilidade que mostram em voltar a casa mesmo quando largados a grandes distâncias. Esta habilidade dos pombos correios nunca foi convenientemente explicada, embora os cientistas tenham avançado com algumas hipóteses. Por exemplo, sabia-se que os pombos são sensíveis aos campos magnéticos, e suspeitava-se que os pombos se guiavam pelo campo magnético terrestre. Ora o facto de se poder fazer algo nem sempre quer dizer que se faça, e pelos vistos os pombos usam um sentido menos misterioso, e mais familiar para nós, para encontrarem o caminho de casa. [... ler mais]
Segundo um artigo de Anna Gagliardo e colegas na revista Journal of Experimental Biology (ref1) os pombos limitam-se a seguir os seus narizes. Numa tradução livre do resumo:A habilidade dos pombos em encontrarem o caminho de casa a partir de locais não familiares, localizados até centenas de km de distância é bem conhecida, mas os mecanismos por trás desta habilidade permanecem controversos. Um mecanismo proposto é baseado na sugestão de que os pombos estão equipados com receptores magnéticos, que permitem a detecção, quer do campo magnético terrestre, quer de anomalias no campo magnético no terreno sobre o qual os pombos voam. Relatos recentes sugerem que estes receptores magnéticos estão localizados no bico superior, onde são inervados pelo ramo oftálmico do nervo trigémeo. Aliás, mostrou-se que este nervo é o mediador da capacidade do pombo em discriminar entre a presença ou ausência de anomalia no campo magnético numa situação condicionada. No estudo presente, contudo, mostramos que um ramo oftálmico do nervo trigémeo não é necessário nem suficiente para um bom desempenho no regresso a casa a partir de locais não familiares, mas que é necessário um nervo olfactivo intacto.
Este resumo é um bocado lacónico, e para compreender melhor como é que os investigadores chegaram a esta conclusão é preciso "mergulhar" no artigo. O que Anna Gagliardo e colegas fizeram foi agarrar em 72 pombos e dividi-los em três grupos quando tinham 5 a 6 meses de idade. A 24 pombos seccionaram o nervo olfactivo, a outros 24 o ramo oftálmico do nervo trigémio (que transporta a informação dos receptores magnéticos), enquanto aos outros 24 fizeram uma operação mas não seccionaram nenhum dos nervos. Metade dos pombos de cada um dos grupos foi largada, 4 dias após a operação, de dois locais com direcções opostas em relação ao seu local de origem e a 50 km de distância do pombal de origem. A restante metade dos pombos foi largada 8 dias após a operação, e dos mesmos locais.
Os resultados não deixaram dúvidas: apenas 4 dos 24 pombos aos quais foram seccionados os nervos olfactivos conseguiram regressar a casa. Nos pombos aos quais se seccionou o nervo trigémeo, ou se fez a operação de controlo, 23 dos 24 pombos em cada grupo encontraram o caminho de casa. Ou seja, os pombos necessitam do seu nervo olfactivo para conseguirem achar o caminho de casa. Este resultado não quer dizer necessariamente que utilizem pistas olfactivas, mas quando combinado com outros estudos em que se taparam as narinas dos pombos, sugere fortemente que esse é o caso.
Para terminar, eis uma foto de um pombo correio adulto, que, embora tenha o seu quê de belo, não exerce sobre mim o mesmo fascínio que os pequenitos.
Ficha técnica
Imagem dos irrestíveis borrachos retirada da Wikimedia Commons, desta página.
Imagem do pombo correio, cortesia de Andreas Trepte, retirada da Wikimedia Commons, desta página.
Referências
(ref1) Gagliardo, A., Ioalè, P., Savini, M. and Wild, J. M. (2006). Having the nerve to home: trigeminal magnetoreceptor versus olfactory mediation of homing in pigeons. J. Exp. Biol. 209,2888 -2892. Laço DOI.
domingo, agosto 27, 2006
Sem lógica para cachorro
 Uma parte importante dos estudos sobre as capacidades mentais dos animais tem a sua origem em estudos não verbais de Piaget sobre as capacidades das crianças para representarem o mundo que as rodeia. Piaget identificou um certo número de estádios no desenvolvimento das crianças, entre eles um que surge entre os 18 e os 24 meses, a partir do qual elas entendem o conceito de deslocamento invisível. Falei numa contribuição anterior sobre as tarefas de deslocamento invisível e de como os cães faziam batota. Para ilustrar esse estudo procurei fotos de cachorros e encontrei tantas que não resisto a mostrar aqui mais uma e usá-la como desculpa para falar de um estudo ligeiramente diferente. [... ler mais]
Uma parte importante dos estudos sobre as capacidades mentais dos animais tem a sua origem em estudos não verbais de Piaget sobre as capacidades das crianças para representarem o mundo que as rodeia. Piaget identificou um certo número de estádios no desenvolvimento das crianças, entre eles um que surge entre os 18 e os 24 meses, a partir do qual elas entendem o conceito de deslocamento invisível. Falei numa contribuição anterior sobre as tarefas de deslocamento invisível e de como os cães faziam batota. Para ilustrar esse estudo procurei fotos de cachorros e encontrei tantas que não resisto a mostrar aqui mais uma e usá-la como desculpa para falar de um estudo ligeiramente diferente. [... ler mais]
Trata-se de um trabalho de 2001 de John S. Watson e colegas publicado na revista Journal of Comparative Psychology (ref1). A motivação para o estudo encontra-se nas primeiras frases do resumo. Numa tradução livre:Pesquisas anteriores sobre a habilidade de resolver a tarefa piagetiana do deslocamento invisível focaram os pré-requisitos para a capacidade representacional. Este estudo examina o pré-requisito da dedução. Tal como noutras tarefas (por exemplo, conservação e transitividade), é difícil distinguir entre comportamento que reflecte inferência lógica do comportamento que reflecte generalização associativa.
Um dos problemas em interpretar resultados de estudos com animais é exactamente esse. Coisas que para nós enquanto seres humanos resultam de pensamento e reflexão, noutros animais podem resultar de mecanismos de associação simples. Para além do exemplo da "batota" dos cães da contribuição anterior, foquei aqui um outro exemplo, aquando da escola das suricatas. Nesse caso os adultos variavam o comportamento em função da idade das crias, dando às mais jovens animais mortos e às mais velhas animais quase intactos. Mas, como os investigadores verificaram, isso não resultava de um julgamento cognitivo avançado por parte dos adultos. Era uma resposta inata ao tipo de vocalização produzida pelas crias, que variava com a idade.
Para testar diferenças a esse nível entre crianças e cães, John Watson e colegas conceberam um teste engenhoso baseado num dispositivo esquematizado na figura abaixo.
Usando o papel da negação na lógica onde uma inferência negativa sobre uma crença aumenta a certeza noutra (por exemplo, um silogismo disjunctivo), cães desconhecedores da tarefa (Canis familiaris; n=19) e crianças de 4 a 6 anos (Homo sapiens; n=24) executaram uma tarefa em que um objecto desejado desaparecia do interior de um dispositivo de transporte depois de passar por detrás de 3 biombos separados.
Os estudiosos da psicologia e do comportamento têm uma certa apetência pelo jargão e pelo encadear de termos complicados, mas o que está aqui descrito é mais simples do que parece. Um objecto que as crianças ou os cães gostavam era colocado dentro de um dispositivo que era em seguida levado ao longo de um trajecto passando por trás de três biombos. Mostrava-se depois o dispositivo (vazio) às crianças ou cães participantes que seguiam o mesmo trajecto em busca do objecto. A parte da "inferência negativa" refere-se ao facto de que, ao verificar que o objecto não se encontra atrás do primeiro e segundo biombos, nós seriamos levados a assumir imediatamente que ele se encontra atrás do terceiro. Ora se as crianças de 4 a 6 anos mostravam evidência de guiarem os seus passos por uma lógica desse tipo, os cães nem por isso:Tal como previsto, as crianças (pela lógica da disjunção negada) tendem a aumentar a sua velocidade para verificar o terceiro biombo após não conseguirem encontrar o objecto atrás dos dois primeiros biombos, enquanto os cães (por extinção associativa) tendem a diminuir a sua velocidade para verificar o terceiro biombo após não encontrarem o objecto atrás dos dois primeiros biombos.
 Apesar do jargão os resultados são fáceis de perceber, e ilustram-se na figura aqui ao lado, onde os tempos nos trajectos do primeiro ao segundo biombo são comparados aos tempos dos trajectos do segundo ao terceiro biombo. Os cães demoravam em geral mais tempo em cada trajecto do que as crianças. mas o importante é que, depois de verem que o objecto não se encontrava nos dois primeiros biombos, as crianças aceleravam o passo do segundo para o terceiro biombo, enquanto os cães faziam o contrário. Este resultado indica que os cães seguem uma estratégia associativa, enquanto as crianças se baseiam, até certo ponto, numa estratégia de inferência lógica quando buscam objectos que desapareceram recentemente. As crianças conseguem antecipar a presença do objecto no terceiro biombo. Os cães não, o que indica entre outras coisas que de facto não compreendem o deslocamento invisível. Este estudo não "prova" que os cães estejam limitados a estas estratégias associativas simples, mas, quando combinado com outros estudos do mesmo tipo, aponta claramente nesse sentido.
Apesar do jargão os resultados são fáceis de perceber, e ilustram-se na figura aqui ao lado, onde os tempos nos trajectos do primeiro ao segundo biombo são comparados aos tempos dos trajectos do segundo ao terceiro biombo. Os cães demoravam em geral mais tempo em cada trajecto do que as crianças. mas o importante é que, depois de verem que o objecto não se encontrava nos dois primeiros biombos, as crianças aceleravam o passo do segundo para o terceiro biombo, enquanto os cães faziam o contrário. Este resultado indica que os cães seguem uma estratégia associativa, enquanto as crianças se baseiam, até certo ponto, numa estratégia de inferência lógica quando buscam objectos que desapareceram recentemente. As crianças conseguem antecipar a presença do objecto no terceiro biombo. Os cães não, o que indica entre outras coisas que de facto não compreendem o deslocamento invisível. Este estudo não "prova" que os cães estejam limitados a estas estratégias associativas simples, mas, quando combinado com outros estudos do mesmo tipo, aponta claramente nesse sentido.
Ficha técnica
Imagem dos cachorros da autoria de Luis Miguel Bugallo Sánchez retirada da Wikimedia Commons, desta página.
Referências
(ref1) John S. Watson, Yorgy Gergely, Vilmos Csanyi, Jozsef Topal, Marta Gacsi, and Zsuzsanna Sarkozi (2001). Distinguishing Logic From Association in the Solution of an Invisible Displacement Task by Children (Homo sapiens) and Dogs (Canis familiaris): Using Negation of Disjunction. Journal of Comparative Psychology, Vol. 115, No. 3, 219-226. Laço DOI.
sexta-feira, agosto 25, 2006
Os cães fazem batota

 Uma das surpresas das tarefas de deslocamento invisível, que referi aqui a propósito dos papagaios cinzentos, é que os cães apresentavam um desempenho acima do que se esperaria se tudo se devesse ao acaso. Isto poderia indicar duas coisas: (1) os cães possuíam capacidades cognitivas avançadas ou (2) o sucesso devia-se a repetição de uma estratégia que por acaso funcionava e não era devido a qualquer tipo de representação mental. Um caso semelhante tinha sido observado em experiências com macacos japoneses, que parececiam compreender o deslocamento invisível, mas afinal se limitavam a escolher a última caixa tocada pelo condutor da experiência. Na verdade, a maioria dos primatas para os quais se tinha relatado algum sucesso nesta tarefa falhou quando se fizeram testes que tentavam controlar algumas desses mecanismos de escolha simples. A excepção foram os grandes símios. Isso levou Emma Collier-Baker e colegas a testarem os cães tendo em atenção esse aspecto. [... ler mais]
Uma das surpresas das tarefas de deslocamento invisível, que referi aqui a propósito dos papagaios cinzentos, é que os cães apresentavam um desempenho acima do que se esperaria se tudo se devesse ao acaso. Isto poderia indicar duas coisas: (1) os cães possuíam capacidades cognitivas avançadas ou (2) o sucesso devia-se a repetição de uma estratégia que por acaso funcionava e não era devido a qualquer tipo de representação mental. Um caso semelhante tinha sido observado em experiências com macacos japoneses, que parececiam compreender o deslocamento invisível, mas afinal se limitavam a escolher a última caixa tocada pelo condutor da experiência. Na verdade, a maioria dos primatas para os quais se tinha relatado algum sucesso nesta tarefa falhou quando se fizeram testes que tentavam controlar algumas desses mecanismos de escolha simples. A excepção foram os grandes símios. Isso levou Emma Collier-Baker e colegas a testarem os cães tendo em atenção esse aspecto. [... ler mais]
Os resultados foram publicados na revista Journal of Comparative Psychology (ref1), e colocam os cães de fora do clube restrito dos animais que compreendem o deslocamento invisível. Numa tradução livre do resumo:Os cães domésticos (Canis familiaris) têm desempenhos acima do acaso em tarefas de deslocamento invisível apesar de mostrarem poucos outros sinais de possuirem as habilidades de representação necessárias. Quatro experiências investigaram como os cães encontram um objecto que foi escondido dentro de uma de três caixas opacas. Os cães concluiram a tarefa com sucesso sob uma variedade de condições de controlo, mas apenas se o dispositivo utilizado para deslocar o objecto fosse colocado adjacente à caixa alvo após o deslocamento. Estes resultados sugerem que o comportamento de busca dos cães era guiado por simples regras associativas e não por uma representação mental da trajectória passada do objecto. Em contraste, a experiência 5 mostrou que, na mesma tarefa, crianças de 18 e 24 meses não revelaram diferenças entre as tentativas em que o dispositivo de deslocamento estava adjacente ou não adjacente à caixa-alvo.
Ou seja, os cães não compreendiam verdadeiramente o processo de transporte e transferência, limitavam-se a procurar na vizinhança do local onde sabiam que o brinquedo que queriam (uma bola) tinha estado. O que guiava as suas buscas era a posição do dispositivo de transporte. Colocando o dispositivo mais perto de outra caixa era nessa que procuravam. As crianças não cometiam esse erro. Esta é uma diferença importante, os cães baseiam-se na percepção imediata que têm do mundo para guiar o seu comportamento, enquanto os seres humanos, mesmo as crianças de dois anos, são capazes de representação secundária, uma capacidade que lhes permite considerar múltiplos modelos do mundo. Este resultado dos cachorros levou Emma Collier-Baker a testar os chimpanzés dentro dos mesmos parâmetros de controlo. Falarei desse estudo numa contribuição futura.
Ficha técnica
Imagem dos cachorros cortesia de Bev Sykes, retirada da Wikimedia Commons, desta página.
Referências
(ref1) Collier-Baker, Emma; Davis, Joanne M.; Suddendorf, Thomas (2004). Do Dogs (Canis familiaris) Understand Invisible Displacement? Journal of Comparative Psychology. 118(4), 421-433. Laço DOI.
quinta-feira, agosto 24, 2006
Sobre a inteligência dos papagaios

 Notei a partir de comentários no Chi vó, non pó, aqui e aqui, acerca de uma notícia sobre a falta de inteligência dos golfinhos, para além de uma certa má vontade dos brasileiros em relação aos sul-africanos, que as pessoas têm um certo apego a esses cetáceos, aos quais atribuem capacidades mentais quase humanas. Resolvi por isso, antes de abordar o estudo sobre os golfinhos, fazer uma pequena série de contribuições sobre questões de inteligência animal. Vou começar não com mamíferos como nós, mas com aves, em particular com o simpático passaroco da figura. Os pagaios cinzentos africanos, Psittacus erithacus, possuem capacidades cognitivas que existem apenas nos seres humanos a partir dos dois anos, e nos grandes símios (chimpanzés e gorilas). [... ler mais]
Notei a partir de comentários no Chi vó, non pó, aqui e aqui, acerca de uma notícia sobre a falta de inteligência dos golfinhos, para além de uma certa má vontade dos brasileiros em relação aos sul-africanos, que as pessoas têm um certo apego a esses cetáceos, aos quais atribuem capacidades mentais quase humanas. Resolvi por isso, antes de abordar o estudo sobre os golfinhos, fazer uma pequena série de contribuições sobre questões de inteligência animal. Vou começar não com mamíferos como nós, mas com aves, em particular com o simpático passaroco da figura. Os pagaios cinzentos africanos, Psittacus erithacus, possuem capacidades cognitivas que existem apenas nos seres humanos a partir dos dois anos, e nos grandes símios (chimpanzés e gorilas). [... ler mais]
A inteligência é um parâmetro difícil de avaliar mesmo nos humanos, quanto mais nos animais. Há no entanto alguns testes simples das capacidades cognitivas que se podem executar. Um deles tem a ver com a permanência dos objectos escondidos. As crianças humanas compreendem o conceito bastante cedo, por volta dos 12 meses de idade. Muitos animais, incluindo chimpanzés, gorilas, orangotangos, várias espécies de macacos, cães, gatos, golfinhos, papagaios, e gralhas, conseguem algo semelhante: percebem que um objecto escondido continua a existir.
Há, no entanto, um limiar acima no desenvolvimento das capacidades cognitivas em que a maioria desses animais falha. As coisas começam a ficar mais interessantes quando se executam tarefas daquilo que se designa por deslocamento invisível. Nesse tipo de testes, geralmente feitos com crianças de tenra idade, um objecto que uma crianças deseja é colocado dentro de um"dispositivo de deslocamento", em geral um pequeno invólucro opaco, e esse dispositivo de deslocamento é então utilizado para transportar o objecto para uma de várias caixas à vista da criança. O investigador coloca então o objecto dentro da caixa escolhida de forma subreptícia sem a criança se aperceber. Em seguida mostra à criança que o objecto que ela queria já não se encontra dentro do dispositivo de deslocamento. Antes dos 18 meses as crianças parecem não perceber o conceito de deslocamento invisível e insistem em procurar dentro do dispositivo de deslocamento apesar dele se encontrar vazio. A partir dos dois anos as crianças vão directamente para a caixa. Segundo Piaget a tarefa de deslocamento invisível é o teste crítico do conceito de permanência do objecto, que marca a transição da criança para a inteligência representacional.
Poucos animais mostram esta capacidade que surge nas crianças entre os 18 a 24 meses. Nos mamíferos não humanos apenas parece existir nos grandes antropóides africanos como os chimpanzés e gorilas. Curiosamente, um estudo de Irene Pepperberg e M.S. Funk na revista Animal Learning & Behavior (ref1) mostra que deverá existir também em várias espécies psitacídeos, um grupo de aves que inclui os papagaios, araras e periquitos. Num outro artigo, na revista Journal of Comparative Psychology (ref2) Irene Pepperberg e colegas fizeram mesmo um estudo mais aprofundado sobre o desenvolvimento dessa capacidade no papagaio-cinzento durante o seu crescimento (ontogenia). Parece surgir bastante cedo. Numa tradução livre do resumo:Os autores avaliaram o desempenho ontogenético de um papagaio cinzento (Psittacus erithacus) em tarefas de permanência de objectos concebidas para crianças humanas. Os testes começaram quando a ave tinha 8 semanas, antes da muda e independência alimentar. Devido ao facto de os papagaios cinzentos adultos perceberem o deslocamento invisível, os autores continuaram os testes semanais até o sujeito completar todos as tarefas da Escala de 1 de I. C. Uzgiris e J. Hunt (1975).
Nessa escala o grau 5 corresponde ao deslocamento visível (esconder simples) e o grau 6 ao deslocamento invisível.Permanência de grau 6 com respeito a essas tarefas surgiu às 22 semanas, após a muda mas antes de começar a alimentar-se sozinho. Embora o papagaio tenha progredido mais rapidamente que outras espécies que foram testadas ontogeneticamente, o sujeito exibiu um patamer de comportamento durante o estudo. Testes adicionais, administrados aos 8 e 12 meses bem como a um papagaio cinzento adulto, mostraram, respectivamente, que estas aves possuem alguma representação de um objecto escondido e que compreendem deslocamentos invisíveis avançados.
Os papagaios cinzentos, apesar de um cérebro relativamente pequeno, parecem estar uns furos acima dos golfinhos e ao nível dos grandes símios. Estes testes terão no entanto que ser repetidos com mais cautela e algum controlo, pois como discutirei aqui proximamente, os cães também pareciam compreender o deslocamento invisível, mas estavam afinal a fazer batota.
Ficha técnica
Imagem do papagaio cinzento no alto da página cortesia de Jason L. Buberal, retirada da Wikimedia Commons, desta página.
Referências
(ref1) Pepperberg IM, Funk MS (1990). Object permanence in four species of psittacine birds: an African grey parrot (Psittacus erithacus), an Illiger mini macaw (Ara maracana), a parakeet (Melopsittacus undulatus), and a cockatiel (Nymphicus hollandicus). Anim Learn Behav 18:97–108
(ref2) Pepperberg IM, Willner MR, Gravitz LB (1997). Development of Piagetian object permanence in a grey parrot (Psittacus erithacus). J Comp Psychol 111:63–75. Laço DOI.
terça-feira, agosto 22, 2006
Os astrónomos são loucos

 Eu já referi aqui por diversas vezes que os astrónomos são criaturas estranhas, embora sempre tenha considerado que os astrónomos planetários, pela sua ligação à Geologia e Exobiologia, eram um pouco mais equilibrados. Pelos vistos são ainda mais estranhos que os outros. Agora arranjaram um imbróglio com os os geólogos por causa da definição de planeta. Para perceber o que se passa é preciso considerar a proposta em inglês, já que em português não há problema. Em Geologia, o mecanismo de formação de rochas ígneas intrusivas é designado por plutonismo (no caso das extrusivas é o vulcanismo). Um exemplo de uma rocha plutónica é o granito. Ora o curioso é que uma intrusão plutónica em inglês é designada por pluton, que segundo a proposta da IAU que discuti aqui, é o nome que se quer dar aos planetas da família de Plutão na versão inglesa. A reacção dos geólogos foi de consternação. [...ler mais]
Eu já referi aqui por diversas vezes que os astrónomos são criaturas estranhas, embora sempre tenha considerado que os astrónomos planetários, pela sua ligação à Geologia e Exobiologia, eram um pouco mais equilibrados. Pelos vistos são ainda mais estranhos que os outros. Agora arranjaram um imbróglio com os os geólogos por causa da definição de planeta. Para perceber o que se passa é preciso considerar a proposta em inglês, já que em português não há problema. Em Geologia, o mecanismo de formação de rochas ígneas intrusivas é designado por plutonismo (no caso das extrusivas é o vulcanismo). Um exemplo de uma rocha plutónica é o granito. Ora o curioso é que uma intrusão plutónica em inglês é designada por pluton, que segundo a proposta da IAU que discuti aqui, é o nome que se quer dar aos planetas da família de Plutão na versão inglesa. A reacção dos geólogos foi de consternação. [...ler mais]
Há um pequeno artigo na Nature News (ref1) sobre esta questão, onde se pode ler uma afirmação fantástica do geólogo Allen Glazner da Universidade do Arizona:Isto é como se os botânicos tivessem encontrado algo entre as árvores e os arbustos e tivessem inventado a palavra "animal" para a descreverem.
As intrusões plutónicas são essencialmente aquilo de que é feita a crosta continental. Não se trata de nada de raro ou pouco comum, daí que seja surpreendente que indivíduos que falam inglês como primeira língua desconheçam a importância do termo pluton. Aliás a Planetologia e a Geologia são parentes muito próximos e o potencial para confusão é imenso. É um bocadinho diferente de Mercúrio planeta e mercúrio metal que raramente aparecerão num contexto que se preste a confusão. Mas o que me fez rir foi a desculpa dos membros da IAU:Owen Gingerich, um astrónomo da Universidade de Harvard em Cambridge, Massachusetts, e presidente da comissão da IAU que criou a definição, diz que eles estavam cientes do seu uso entre os geólogos, mas que não se tinham apercebida da sua importância para o ramo. Num email para news@nature.com Gingerich escreveu "Como o termo não aparece nos correctores ortográficos do MS Word ou do WordPerfect pensámos que não era assim tão usual".
O MS Word como autoridade científica! Pelos visto é o Bill Gates que decide se um termo é cientificamente relevante ou não.
Adenda: a IAU substituiu, na proposta a votação, pluton por plutonian. Em português também faz um bocado mais sentido, pluton seria traduzido por plutónio. Assim ficamos com plutoniano. Mais adenda: a proposta do plutonian foi recusada.
Ficha técnica
Fotografia de granito rosa em França (Bretagne Côtes d'Armor) cortesia de Patrick Giraud tirada da Wikimedia Commons, desta página.
Referências
(ref1) Geoff Brumfiel (2006). Plutons, planets and dwarves Geologists and astronomers wrangle over words.Laço DOI.
A mandíbula mais rápida do oeste

 Por vezes a Natureza surpreende-nos. Mesmo um animal tão comum como uma formiga pode mostrar algo de que não se está à espera. Por vezes uma estrutura que tendemos a associar a uma função pode servir para algo completamente diferente, e ser extremamente eficiente a desempenhar essa outra função. As mandíbulas das formigas servem para morder e a função delas é essencialmente essa, certo? Bem, nem por isso. Pelo menos não para a Odontomachus bauri, uma espécie de formigas das Selvas da Costa Rica com mandíbulas realmente impressionantes. Capazes de uma abertura de cerca de 180 graus, essas mandíbulas são adequadas aos hábitos predatórios destas criaturas com apenas alguns milímetros de comprimento. O que é extraordinário é que as formigas utlizam as mandíbulas para escaparem a situações de perigo, não confrontando os adversários mas fugindo deles a toda a velocidade. De facto, estas mandíbulas permitem a essas formigas feitos que envergonhariam qualquer atleta humano. E não estou a falar apenas de levantamento de pesos ou outras coisas óbvias. Estou a falar de velocidade, e saltos em altura e comprimento. O que vocês diriam, nestes tempos em que o petróleo está tão caro, se fossem capazes de levantar vôo a velocidades elevadas com um simples bater de dentes? Pois bem, a Odontomachus bauri é capaz disso, e podemos apreciar esse desempenho em filme. [... ler mais]
Por vezes a Natureza surpreende-nos. Mesmo um animal tão comum como uma formiga pode mostrar algo de que não se está à espera. Por vezes uma estrutura que tendemos a associar a uma função pode servir para algo completamente diferente, e ser extremamente eficiente a desempenhar essa outra função. As mandíbulas das formigas servem para morder e a função delas é essencialmente essa, certo? Bem, nem por isso. Pelo menos não para a Odontomachus bauri, uma espécie de formigas das Selvas da Costa Rica com mandíbulas realmente impressionantes. Capazes de uma abertura de cerca de 180 graus, essas mandíbulas são adequadas aos hábitos predatórios destas criaturas com apenas alguns milímetros de comprimento. O que é extraordinário é que as formigas utlizam as mandíbulas para escaparem a situações de perigo, não confrontando os adversários mas fugindo deles a toda a velocidade. De facto, estas mandíbulas permitem a essas formigas feitos que envergonhariam qualquer atleta humano. E não estou a falar apenas de levantamento de pesos ou outras coisas óbvias. Estou a falar de velocidade, e saltos em altura e comprimento. O que vocês diriam, nestes tempos em que o petróleo está tão caro, se fossem capazes de levantar vôo a velocidades elevadas com um simples bater de dentes? Pois bem, a Odontomachus bauri é capaz disso, e podemos apreciar esse desempenho em filme. [... ler mais]
As proezas da Odontomachus bauri foram analisadas por S.N. Patek e colegas, que apresentam os resultados nos Proceedings of the National Academy of Sciences USA (ref1). Numa tradução livre do resumo:Movimentos extremos nos animais estão usualmente associados com um único comportamento, de elevado desempenho. Contudo, os fechares de mandíbulas notavelmente rápidos da formiga Odontomachus bauri podem resultar em variados resultados funcionais. Investigamos aqui a biomcânica dos fechares de mandíbula na O. bauri e verificamos que os movimentos de mandíbula extremos servem duas funções distintas: predação e propulsão. Durante ataques predatórios, as mandíbulas da O. bauri fecham a velocidades que vão de 35 a 64 metros por segundo, com uma duração média de 0.13 ms, ultrapassando de longe a velocidade de outros apêndices balísticos documentados no reino animal.
Os autores colocam um filme como material suplementar de apoio ao artigo, que pode ser visto aqui. Este filme foi produzido com uma câmara que filmava a 50,000 imagens por segundo (!) e é mostrado a 30 imagens por segundo. Para terem uma ideia do que está em jogo, é como se conseguissemos produzir cerca de 20 toneladas de força com o bater dos nossos dentes:As elevadas velocidades das mandíbulas auxiliam na captura de presas, enquanto as acelerações extremas resultam em forças instantâneas de bater das mandíbulas que podem exceder 300 vezes o peso da formiga.
Isto permite algo de verdadeiramente impressionante:Como consequência, uma batida de mandíbulas da O. bauri dirigida contra o substrato produz suficiente poder proulsor para lançar a formiga no ar. Mudar a orientação da cabeça e da superfície permite à O. bauri utilizar o seu mecanismo mandibular para capturar presas, expulsar intrusos, ou saltar para a segurança. Esta utilização de um único sistema mecânico simples para gerar um conjunto de comportamentos profundamente diferentes oferece uma nova visão das origens morfológicas das novidades na locomoção e na procura de alimento.
O verdadeiro poder do apertar de mandíbulas para a fuga pode apreciado em mais um filme, desta vez filmado a 3,000 imagens por segundo, e mostrado a 30 imagens segundo, de um espectacular "salto em altura" defensivo. Mesmo em câmara lenta a velocidade é extraordinária e bater com a cabeça no chão daquela maneira deve doer. Outro filme mostra aqui mais um exemplo de um salto em altura. Confesso que as quedas me impressionam. E reparem que a acção real é cerca de 100 vezes mais rápida! Eis aqui algumas imagens para aqueles que não dispõem de ligação rápida à internet ou que por qualquer razão não conseguem ver os filmes.
O número no canto superior esquerdo de cada uma das imagens é o número do fototgrama no filme (lembrem-se que 3,000 significa um segundo). As possibilidades de utilização das mandíbulas para a fuga são igualmente impressionantes se utilizadas no "salto em comprimento". Este filme permite compreender como o lançamento se processa. Outro filme que se mostra aqui mostra o alcance de um destes saltos. Alguns fotogramas para aqueles que não possam ou prefiram não ver o filme:
Verdadeiramente impressionante, embora continue a achar que têm que trabalhar mais as aterragens. Se não vos parece nada de mais, notem só que à escala humana o salto em altura destas formigas corresponde a um salto acima dos 13 metros, e o salto em comprimento mais de 40 metros. Já agora, uma tão notável criatura merece uma fotografia de corpo inteiro em alta resolução.
Ficha técnica
Imagem de Odontomachus bauri no início da contribuição tirada por C. Richart, obtida nesta página.
Os filmes e os fotogramas mostrados podem obter-se a partir do laço indicado na ref1 abaixo.
Referências
(ref1) S. N. Patek, J. E. Baio, B. L. Fisher, and A. V. Suarez (2006). Multifunctionality and mechanical origins: Ballistic jaw propulsion in trap-jaw ants. PNAS. Laço DOI.
sábado, agosto 19, 2006
As lentes virtuais das cobras

 Os crotalíneos como esta cascavél, de nome científico Crotalus willardi obscurus, são uma família de cobras que apresentam duas cavidades, entre cada uma das narinas e cada um dos olhos, onde se encontram membranas sensíveis à radiação infravermelha, isto é ao calor. Essa pequena fossa vê-se claramente na imagem. Estes termoreceptores permitem às serpentes produzirem das suas presas imagens no infravermelho. Ora, não possuindo uma lente, as imagens seriam em princípio semelhantes às que se obtêm numa câmara que produz fotografia estenopeica. [... ler mais]
Os crotalíneos como esta cascavél, de nome científico Crotalus willardi obscurus, são uma família de cobras que apresentam duas cavidades, entre cada uma das narinas e cada um dos olhos, onde se encontram membranas sensíveis à radiação infravermelha, isto é ao calor. Essa pequena fossa vê-se claramente na imagem. Estes termoreceptores permitem às serpentes produzirem das suas presas imagens no infravermelho. Ora, não possuindo uma lente, as imagens seriam em princípio semelhantes às que se obtêm numa câmara que produz fotografia estenopeica. [... ler mais]
Neste tipo de fotografia, uma abertura muito pequena, tipicamente com menos de meio mílimetro de diâmetro, deixa passar a luz para dentro de uma caixa onde produz uma imagem invertida. Em inglês este tipo de câmaras são designadas por pinholes, ou seja buraco de alfinete. As caixas estenopeicas são populares, por exemplo, para ver eclipses por projecção da imagem. O problema é que, no caso da serpente, para conseguir infravermelhos suficientes para produzir uma imagem em tempo útil, a abertura tem que ser bastante maior que um buraco de alfinete, e a qualidade das imagens deveria ser muito má. Mas na verdade não é isso que se passa, e Andreas B. Sichert e colegas num artigo na Physical Review Letters (ref1) explicam porquê.
Se os termoreceptores funcionassem como uma câmara estenopeica clássica, confrontada com o coelho na imagem abaixo à esquerda, da autoria do pintor Albrecht Durer, uma cascavel veria o borrão que se encontra à direita.
O desempenho de um tal sistema seria muito pobre. Contudo, na natureza, as cobras comportam-se como se conseguissem discriminar detalhes muitíssimos menores. Afinal, têm que ser capazes de acertar num rato a uma distância da ordem de um metro. As cobras, pelos vistos, terão um elaborado sistema de tratamento de imagens implementado no seu cérebro. O problema é que as serpentes têm "pouco" cérebro. O que os autores do artigo investigaram foi exactamente quão grande precisa de ser o número de neurónios para produzir uma imagem decente no infravermelho. Aparentemente pode ser bastante pequeno. Numa tradução livre do resumo do artigo:Dois grupos de cobras possum um sistema de detecção no infravermelho que é utilizado para criar uma imagem de calor do seu ambiente. Nesta Letter apresentamos um modelo de reconstrução explícita, a "lente virtual", que explica como a serpente pode ultrapassar as limitações ópticas de uma câmara estenopeica de grande abertura, e como as propriedades dos campos receptores na membrana sensível ao infravermelho podem explicar o desempenho comportamental deste sistema sensorial. O nosso modelo explora a qualidade óptica do sistema de infravermelhos mostrando em detalhe como se pode criar uma representação funcional das propriedades térmicas do ambiente. O modelo é fácil de implementar do ponto de vista neuronal e está em bom acordo com os dados neuronais, fisiológicos e comportamentais, disponíveis sobre o sistema de infravermelho das cobras.
O que os cientistas fizeram foi algo indirecto. O que eles mostraram não foi exactamente o que as serpentes fazem, mas que um animal com um cérebro tão pequeno como uma cobra poderia implementar apesar de tudo um algoritmo muito eficiente para o tratamento de imagens. Os autores assumem que os detectores de calor em cada uma das fossas termoreceptoras fazem "disparar" um sinal num neurónio. A taxa de "disparo" varia com os sinais sentidos nos outros receptores, e, usando um algoritmo bastante simples para "afinar" a interacção dos cerca de 2,000 sensores de calores presentes nas membranas, Andreas B. Sichert e colegas conseguiram imagens de um detalhe impressionante. A imagem abaixo mostra como a mesma imagem do coelho seria vista por uma cascavel virtual. É notável, consegue já distinguir-se a forma do animal.
Este processamento de imagem exige apenas uns poucos milhares de neurónios. É óbvio que as cobras utilizarão um algoritmo algo diferente do desenvolvido por Andreas B. Sichert e colegas. O que o trabalho mostra é que a tarefa está ao alcance das capacidades mentais de uma cobra. Não é preciso uma lente para produzir imagens de boa qualidade no infravermelho quando se tem um cérebro, por muito pequeno que seja. As aplicações prácticas deste estudo são óbvias e vão para além da compreensão da visão das cobras.
Ficha técnica
Foto de Crotalus willardi obscurus tirada por Robert S. Simmons da USFWS, pode ser encontrada na Wikimedia Commons, aqui.
Referências
(ref1) Andreas B. Sichert, Paul Friedel, and J. Leo van Hemmen (2006). Snake's Perspective on Heat: Reconstruction of Input Using an Imperfect Detection System. Phys. Rev. Lett. 97, 068105. Laço DOI.
sexta-feira, agosto 18, 2006
Blogues e a dificuldade em divulgar temas de Biologia
 Na Roda de Ciência, o tema do mês é a questão do jornalismo científico, divulgação científica, e o papel dos cientistas nessa divulgação. Embora a população leitora de blogues seja pequena, não é tão pequena como isso, e dá para retirar algumas conclusões sobre o real interesse da população pelos temas científicos. Notei que alguns são mais populares que outros. Os meus leitores parecem não se interessar muito por Biologia, o que me incomoda um pouco, pois cerca de 70% das minhas contribuições são sobre esse tema.
Na Roda de Ciência, o tema do mês é a questão do jornalismo científico, divulgação científica, e o papel dos cientistas nessa divulgação. Embora a população leitora de blogues seja pequena, não é tão pequena como isso, e dá para retirar algumas conclusões sobre o real interesse da população pelos temas científicos. Notei que alguns são mais populares que outros. Os meus leitores parecem não se interessar muito por Biologia, o que me incomoda um pouco, pois cerca de 70% das minhas contribuições são sobre esse tema.
[... ler mais]
Antes de analisar os problemas de popularidade dos vários temas, é importante não escamotear as diferenças enormes que existem entre um blogue e os meios de divulgação tradicionais, como colunas em jornais, palestras, ou mesmo as páginas da Internet num formato mais tradicional. Num blogue fala-se sobretudo daquilo de que se gosta. Há, por exemplo, blogues de cientistas que começaram com a intenção de falar um pouco de ciência mas resvalaram completamente para temas políticos/artísticos. O caso mais óbvio, que apesar de tudo mantenho na lista de Ciência em Português, é o Linha dos Nodos. Por outro lado, num blog pode falar-se de tudo um pouco, incluindo aspectos da vida pessoal, e pode dizer-se de alguns blogs (Osame cadê você?) que têm mesmo um carácter intimista.
O Cais de Gaia, ao contrário da maioria dos blogues na roda de ciência, é um blogue de não especialista, de carácter jornalístico. A génese seguiu um trajecto um pouco ao contrário de outros blogues. Por estranho que possa parecer, na primeira versão era um blogue sobre futebol. A passagem à ciência foi apesar de tudo natural, pois se existe uma ampla escolha de blogues para satisfazer as minhas necessidades futebolísticas, não abundam na internet locais escritos em Português que se situem a um nível intermédio entre o blogue de especialista e o blogue de entusiasta. Como blogue é algo eclético, qualquer artigo que leia e ache interessante falo nele. Uma das coisas que mais me surpreendeu foi a dificuldade de captar público com os temas de Biologia, e mesmo Geologia, que não tenham que ver com Astronomia. É curioso mas páginas sobre as novas Luas de Plutão, o tal objecto alcunhado de Xena maior que plutão, o asteróide Apófis, géiseres em Encélado, Xanadu em Titã, ou mais recentemente a tal coisa de Marte ser visto com o tamanho aparente da Lua, receberam milhares de visitas em poucos dias. Nenhuma das minhas páginas sobre Biologia ou temas afins como Paleontologia recebeu muito para além da centena ou poucas centenas de visitantes. Estamos a falar de uma diferença de mais do que uma ordem de grandeza.
A explicação poderia ser algum defeito na minha escrita, mas a impressão que tenho é que os blogues de biologia em português têm mesmo pouca procura. Este desinteresse é algo paradoxal quando se pensa que o blog de ciência mais popular nos EUA é o pharyngula, exactamente sobre temas ligados à Biologia e evolução. Será que o sucesso que têm nos EUA se deve sobretudo ao problema do criacionismo, que não existe em Portugal e no Brasil? Será que tem a ver com diferentes níveis de exposição nos meios de comunicação tradicionais, e a blogosfera ainda se ressente disso?
COMENTÁRIOS (via roda de ciência)
![]()
quarta-feira, agosto 16, 2006
Abaixo da superfície do Etna

 Esta soberba imagem tirada do espaço, a partir da Estação Espacial Internacional mostra uma visão da erupção no dia 30 de Outubro de 2002, cuja pluma de gás e cinza se eleva a 9000 metros acima do nível do mar e se curva no horizonte levando cinza até à Africa do Norte, a centenas de km de distância. Aliás, a costa africana é mesmo visível lá longe no horizonte, no topo da imagem. Esta fotografia é impressionante pois mostra não só a estrutura vertical da pluma mas também a sua extensão. Uns poucos minutos depois a tripulação mudou as lentes da câmara e obteve um panorama com uma abertura angular menor mas igualmente impressionante.[...ler mais]
Esta soberba imagem tirada do espaço, a partir da Estação Espacial Internacional mostra uma visão da erupção no dia 30 de Outubro de 2002, cuja pluma de gás e cinza se eleva a 9000 metros acima do nível do mar e se curva no horizonte levando cinza até à Africa do Norte, a centenas de km de distância. Aliás, a costa africana é mesmo visível lá longe no horizonte, no topo da imagem. Esta fotografia é impressionante pois mostra não só a estrutura vertical da pluma mas também a sua extensão. Uns poucos minutos depois a tripulação mudou as lentes da câmara e obteve um panorama com uma abertura angular menor mas igualmente impressionante.[...ler mais] A estrutura vertical da pluma, que se eleva a 6400 metros acima do topo da cratera do vulcão, pode ser apreciada nestas imagens espaciais de uma forma que não é possível mesmo com fotografias em aviões.
A estrutura vertical da pluma, que se eleva a 6400 metros acima do topo da cratera do vulcão, pode ser apreciada nestas imagens espaciais de uma forma que não é possível mesmo com fotografias em aviões.
O artigo que me permitiu voltar a este fascinante tema, de Patanè e colegas, na revista Science (ref1), discute a inversão de dados de sismologia por forma a construir um modelo do que se passa por baixo da superfície de um vulcão durante uma erupção. Antes de passar ao tradicional resumo do artigo convém introduzir alguns dos diferentes tipos de ondas que resultam de abalos sísmicos. As ondas sísmicas mais importantes são as ondas compressivas (P), ondas de cisalhamento (S), ondas de Rayleigh (R), e ondas de Love (L). Para o artigo de Patanè e colegas as que interessam são as P e S.
As ondas P não são mais que as vulgares ondas sonoras, que se propagam no ar a cerca de 300 metros por segundo e na crosta terrestre a valores entre 5 a 7 km por segundo. Chamam-se ondas compressivas porque alternam compressões e dilatações no sentido de propagação da onda. Se isto parece pouco claro, aconselho vivamente a animação que se pode ver carregando na imagem abaixo, cortesia de L. Braile (ver ficha técnica). As ondas S ou de cisalhamento, são mais lentas que as ondas P, com velocidades típicas na crosta de 3 a 4 km por segundo. Estas ondas exigem um substrato sólido, não se propagam através de fluídos como o ar ou água líquida. Caracterizam-se, tal como o nome indica, por oscilações perpendiculares ao sentido de propagação da onda. Se isto parece algo obscuro, aconselho a visualização de mais uma animação de L. Braile carregando na imagem abaixo.
As ondas S ou de cisalhamento, são mais lentas que as ondas P, com velocidades típicas na crosta de 3 a 4 km por segundo. Estas ondas exigem um substrato sólido, não se propagam através de fluídos como o ar ou água líquida. Caracterizam-se, tal como o nome indica, por oscilações perpendiculares ao sentido de propagação da onda. Se isto parece algo obscuro, aconselho a visualização de mais uma animação de L. Braile carregando na imagem abaixo. Passemos então ao resumo do artigo de Patanè e colegas.
Passemos então ao resumo do artigo de Patanè e colegas.As continuadas actividades vulcânica e sísmica no Monte Etna fazem deste vulcão um laboratório importante para estudos sísmicos e geofísicos. Utilizámos repetições de tomografia tridimensional para detectar variações nos parâmetros elásticos durante diferentes ciclos vulcânicos, antes e depois da erupção no flanco que ocorreu de Outubro de 2002 a Janeiro de 2003. Foram revelados valores bem definidos baixo e anómalos dos quocientes das velocidades das ondas P e S.
O que os autores fizeram foi utilizar uma vasta rede de sismógrafos na Ilha da Sicília para construir um mapa das ondas sísmicas em profundidade para vários valores da latitude e longitude, isto é puderam construir mapas tridimensionais da velocidade em toda a zona em redor do vulcão. Estes mapas tridimensionais foram construídos em períodos diferentes, daí a referência a tomografia quadridimensional. Ora o quociente entre a velocidade das ondas P e S varia com as características do material que atravessam, e foi esse parâmetro que os autores utilizaram para compreender o que sucedia nas entranhas do vulcão durante a erupção.
A figura abaixo mostra alguns cortes bidimensionais feitos em três intervalos de tempo diferentes e a latitudes diferentes. Todos os gráficos mostram o quociente entre a velocidade das ondas P e S em função da altitude e de longitude. A escala de cores no topo da imagem dá os valores do quociente que vão desde 1.6 para o vermelho a 2.1 para o violeta. Os contornos branco e vermelho delimitam em cada gráfico regiões onde o quociente pôde ser determinado com grande confiança. Os gráficos da esquerda (A) e centro (B) indicam períodos anteriores à erupção, enquanto os gráficos da coluna da direita (C) são referentes a um período durante a erupção. Se repararem com atenção nos gráficos C podem verificar que há uma pequena mancha vermelha que corresponde a um local onde o quociente entre a velocidade das ondas P e S é muito baixo. Trabalhos anteriores tinham mostrado que isso acontece, por exemplo, se o material for rico em material gasoso, embora haja outras possibilidades para esse valor anómalo, que precisam de ser testadas com informação complementar. Continuando com o resumo:
Se repararem com atenção nos gráficos C podem verificar que há uma pequena mancha vermelha que corresponde a um local onde o quociente entre a velocidade das ondas P e S é muito baixo. Trabalhos anteriores tinham mostrado que isso acontece, por exemplo, se o material for rico em material gasoso, embora haja outras possibilidades para esse valor anómalo, que precisam de ser testadas com informação complementar. Continuando com o resumo: Ausentes durante o período pré-eruptivo, as anomalias traçam a intrusão de magma basáltico rico em voláteis (>4 em percentagem do peso), a maior parte do qual se elevou apenas uns poucos meses antes do início da erupção. As mudanças temporais observadas nas anomalias da velocidade sugerem que a tomografia quadridimensional fornece uma base para mais eficientes monitorização dos vulcões e previsões a curto e médio prazos da actividade explosiva.
Tal como indicado na parte que destaquei a negrito, esta técnica tem um potencial enorme para os esforços de previsão, embora por enquanto o tipo de cobertura por sismógrafos que pôde ser utilizado aqui não exista para a grande maioria dos vulcões activos. O estudo de Patanè e colegas é discutido num outro artigo no mesmo volume da Science (ref2) da autoria de Gillian R. Foulger. Nesse artigo, para além de uma curta discussão dos princípios subjacentes à técnica utilizada e referência a estudos anteriores, a autora discute alguns dos cuidados a ter quando se interpretam estes resultados. Não vou entrar em detalhes pois esta contribuição já vai longa, deixo apenas o sugestivo título desse artigo:Geofísica: em direcção à tecnologia dos supervulcões.
Ficha técnica
As animações apresentadas, e exemplos também das ondas de Love e Rayleigh, podem ser encontradas nas páginas de L. Braile na Universidade de Purdue, Seismic Wave Demonstrations and Animations.
Referência
(ref1) D. Patanè, G. Barberi, O. Cocina, P. De Gori, C. Chiarabba (2006). Time-Resolved Seismic Tomography Detects Magma Intrusions at Mount Etna. Science Vol. 313. no. 5788, pp. 821 - 823. Laço DOI.
(ref2) Gillian R. Foulger (2006). GEOPHYSICS: Toward "Supervolcano" Technology. Science Vol. 313. no. 5788, pp. 768 - 769. Laço DOI.
terça-feira, agosto 15, 2006
Oceanos de bactérias

 Eu já falei aqui no Censo da Vida Marinha, em particular a propósito de pterópodes. Mas o Censo da Vida Marinha não se interessa apenas por caracóis voadores, investiga também criaturas muitíssimo menores. Existe mesmo aquilo que se designa por Censo Internacional dos Micróbios Marinhos, que tem feito esforços para aumentar a eficiência dos processos que avaliam a riqueza taxonómica dos oceanos. A situação nos últimos anos progrediu rapidamente devido a técnicas que permitem analisar directamente o ADN de uma amostra, sem ser preciso cultivar os diferentes microrganismos em laboratório como no passado. Esse foi um grande avanço, pois apenas se consegue cultivar em laboratório uma ínfima fracção dos micróbios marinhos. Contudo, identificar o número de tipos de micróbios continua a não ser uma tarefa fácil. [... ler mais]
Eu já falei aqui no Censo da Vida Marinha, em particular a propósito de pterópodes. Mas o Censo da Vida Marinha não se interessa apenas por caracóis voadores, investiga também criaturas muitíssimo menores. Existe mesmo aquilo que se designa por Censo Internacional dos Micróbios Marinhos, que tem feito esforços para aumentar a eficiência dos processos que avaliam a riqueza taxonómica dos oceanos. A situação nos últimos anos progrediu rapidamente devido a técnicas que permitem analisar directamente o ADN de uma amostra, sem ser preciso cultivar os diferentes microrganismos em laboratório como no passado. Esse foi um grande avanço, pois apenas se consegue cultivar em laboratório uma ínfima fracção dos micróbios marinhos. Contudo, identificar o número de tipos de micróbios continua a não ser uma tarefa fácil. [... ler mais]
Num mililitro de água do mar podem existir de 100 mil a um milhão de microrganismos, mas esse número é em grande parte devido a uns poucos tipos de micróbios. Daí que, enquanto esses tipos mais comuns são fáceis de identificar mesmo com amostras pequenas, identificar as estirpes mais raras exige que se investiguem volumes muito maiores do que aqueles que se descrevem na literatura. Os trabalhos publicados referem desde uma poucas centenas até um máximo de 3,000 tipos de micróbios por litro de água mar. Ora uma investigação recente mostra que estes valores pecam claramente por defeito.
Fruto do trabalho desenvolvido pelo Censo Internacional dos Micróbios Marinhos, Mitchell L. Sogin e colegas relatam, num artigo nos Proceedings of the National Academy of Sciences USA (ref1), resultados relativos à abundância e número de estirpes de bactérias que se podem encontrar na água das profundezas dos oceanos. Vamos então ao artigo, e nada melhor do que começar com alguns números que os autores referem na introdução:Os oceanos do mundo pululam com formas de vida microscópicas. Contagens nominais de >105 células por mililitro na superfície da água do mar prevêm que os oceanos acolhem 3.6 x 1029 células microbiais com conteúdo total de carbono de aproximadamente 3 x 1017 g. Comunidades de bactérias, arquebactérias, protistas, e fungos unicelulares respondem pela maior parte da biomassa dos oceanos. Estas fábricas microscópicas são responsáveis por 98% da produção primária de energia e medeiam todos os ciclos biogeoquímicos nos oceanos.
O 3 seguido de 17 zeros pode não parecer impressionante, mas reparemos que se admitirmos um peso médio de cerca de 60 kg por pessoa, dos quais cerca de 20% é carbono, os cerca de 6,000 milhões de seres humanos no nosso planeta contêm "apenas" cerca 7.2x 1013 g de carbono, ou seja 4,000 vezes menos.
Antes de introduzir o habitual resumo, convém salientar um ponto importante. Nos animais, uma espécie é em geral um grupo de indivíduos que em população produzem descendentes férteis cujo material genético é uma combinação do material genético dos progenitores. A chave aqui é o baralhar de material genético e a possibilidade de fertilização cruzada. Uma espécie animal pode assim ser encarada como uma unidade evolutiva. Mas mesmo nos animais há excepções, naqueles que se reproduzem por partenogénese, isto é, de forma assexuada, e são conjuntos de clones. Neste caso, falar em "espécie" tem um valor menos óbvio do ponto de vista conceptual.
O conceito de espécie também não faz muito sentido em micróbios como as bactérias, que se reproduzem assexuadamente. Embora nas bactérias possa haver troca de material genético entre indivíduos diferentes, através de um processo chamado transferência lateral, esse processo não é parte determinante ou necessária do processo reprodutivo. Sendo assim faz muito mais sentido não falar em espécies mas utilizar um indicador de semelhança genética, que é colocado num certo limite e nos dá uma ideia da variabilidade genética na amostra de micróbios. Passando então ao resumo:A evolução dos micróbios marinhos ao longo de milhares de milhões de anos prevê que a composição das comunidades microbianas deveria ser muito superior às estimativas de uns poucos milhares de tipos diferentes de micróbios por litro de água do mar. Ao adoptar uma estratégia paralela de identificação maciça de marcadores, mostramos que as comunidades de bactérias das águas profundas do mar do Atlântico Norte e fontes hidrotermais dos fundos oceânicos são uma ou duas ordens de grandeza mais complexas do que anteriormente relatado para quaisquer ambientes microbianos.
Os números no artigo são impressionantes. Os autores utilizam métodos estatísticos, as chamadas curvas de rarefação, para estimar quantas estirpes poderão existir numa dada amostra usando critérios de diferença de 3%, 5% e 10%. De acordo com o critério mais alargado (3%), que os autores utilizam para indentificar o "tipo" de micróbio, mas apesar de tudo suficientemente amplo para evitar os erros de identificação, numa das amostras existiriam mais de 23,000 estirpes distintas de micróbios num litro de água. Mesmo com um critério bastante mais restritivo (10%), que indica o número de estirpes muito divergentes, essa amostra conteria pelo menos 8,000.Um número relativamente pequeno de diferentes populações domina todas as amostras, mas as centenas de populações de baixa abundância respondem pela maior parte da diversidade filogenética. Esta "biosfera rarefeita" é muito antiga e pode representar uma fonte quanse inesgotável de inovações genómicas. Os membros da biosfera rarefeita apresentam uma grande divergência entre si e, em diferentes períodos na história da Terra, podem ter tido um impacto profundo no modelar dos processos planetários.
A afirmação com o ênfase a negrito significa que embora raras agora nada impede que algumas destas estirpes possam ter sido as formas dominantes durante períodos em que as condições nos oceanos eram diferentes, nem que possam vir a ser dominantes num futuro mais ou menos próximo, se as coisas se alterarem. As bactérias e arquebactérias marinhas, com um número de estirpes agora inflacionado a qualquer coisa entre 5 a 10 milhões, representam cerca 90% da biomassa dos oceanos. Os mares da Terra pertencem de facto às bactérias.
Ficha técnica
Esta contribuição beneficiou das discussões no Deep Sea News.
O comunicado de imprensa do Censo da Vida Marinha, de onde retirei a imagem das bactérias que usei na composição no alto da contribuição, pode ser encontrado aqui.
Referências
(ref1) Mitchell L. Sogin, Hilary G. Morrison, Julie A. Huber, David Mark Welch, Susan M. Huse, Phillip R. Neal, Jesus M. Arrieta, and Gerhard J. Herndl (2006). Microbial diversity in the deep sea and the underexplored "rare biosphere". Proc. Natl. Acad. Sci. USA, Laço DOI
segunda-feira, agosto 14, 2006
As coisas que os químicos podem fazer na cozinha
 Eis algo que descobri através do blog da GrrlScientist, Living the Scientific Life (Scientist, Interrupted).Se quer saber o acontece quando se juntam 200 litros de coca-cola de dieta e 500 pastilhas de Mentos, eis a sua oportunidade. Ainda por cima pode ver tudo em filme (exige ligação rápida e paciência, são cerca de 12 Mbytes). [... ler mais]
Eis algo que descobri através do blog da GrrlScientist, Living the Scientific Life (Scientist, Interrupted).Se quer saber o acontece quando se juntam 200 litros de coca-cola de dieta e 500 pastilhas de Mentos, eis a sua oportunidade. Ainda por cima pode ver tudo em filme (exige ligação rápida e paciência, são cerca de 12 Mbytes). [... ler mais]
Já agora, os cientistas pensam que a causa dos géiseres é física e não química. As rugosidades e poros na pastilha dos mentos permitem que se formem quantidades colossais de bolhas muito depressa. Embora isto não queira dizer que se corra o risco de "rebentar", caso se beba coca-cola e se comam Mentos ao mesmo tempo, os autores do vídeo alertam as pessoas para que não testem os limites dos respectivos estômagos.
Adenda: fui demasiado rápido ao referir o que os cientistas pensam. Há quem defenda um "modelo químico". O Douglas Adamoski Meira fez um trabalho de pesquisa bastante exaustivo e explica as várias soluções de forma bastante detalhada no seu blog Leituras para um ócio produtivo.
Ficha técnica
O filme e uma explicação de como funciona (em inglês) cortesia de EepyBird.com
O elefante que caça pinguins em terra

 Depois de anteriormente ter falado de elefantes que caçam borboletas no mar, hoje falo de elefantes que caçam pinguins em terra. A imagem mostra um elefante-marinho, de seu nome científico Mirounga leonina, uma espécie em que os machos durante a época de reprodução se entregam a violentas batalhas pelo controlo de um território e de um harém de fêmeas. Este macho não teve tanta sorte e dedicou-se durante esse período a uma actividade com o seu quê de surpreendente. Se repararem com atenção, à esquerda da cabeça do elefante-marinho, junto à pedra na parte inferior da imagem, veêm uma espécie de pequena trouxa felpuda. Trata-se dos restos mortais de um pinguim magalhânico, Spheniscus magellanicus, e a foca foi a culpada do seu triste fim. Mas não se trata de uma simples história de predador contra presa. Os motivos deste caçador, mais que a fome, parecem ter sido o tédio, a frustração ou a ira. [... ler mais]
Depois de anteriormente ter falado de elefantes que caçam borboletas no mar, hoje falo de elefantes que caçam pinguins em terra. A imagem mostra um elefante-marinho, de seu nome científico Mirounga leonina, uma espécie em que os machos durante a época de reprodução se entregam a violentas batalhas pelo controlo de um território e de um harém de fêmeas. Este macho não teve tanta sorte e dedicou-se durante esse período a uma actividade com o seu quê de surpreendente. Se repararem com atenção, à esquerda da cabeça do elefante-marinho, junto à pedra na parte inferior da imagem, veêm uma espécie de pequena trouxa felpuda. Trata-se dos restos mortais de um pinguim magalhânico, Spheniscus magellanicus, e a foca foi a culpada do seu triste fim. Mas não se trata de uma simples história de predador contra presa. Os motivos deste caçador, mais que a fome, parecem ter sido o tédio, a frustração ou a ira. [... ler mais]
Pelo menos essas são as conclusões de um interessante artigo de J. Alan Clark e P. Dee Boersma que encontrei no Marine Mammal Science (ref1). Numa tradução livre do resumo:A predação de pinguins por pinípedes no mar está bem documentada. Contudo, a predação de pinguins por pinípedes em terra é rara. Durante o período de Setembro a Novembro de 1997-1999, uma foca elefante macho (Mirounga leonina) matou pelo menos 88 pinguins magalhânicos (Spheniscus magellanicus) na Reserva Provincial de at Punta Tombo (44 02 S, 65 11 W) em Chubut, na Argentina, tendo um impacto significativo na colónia.
A vida dos pinguins não é fácil, perseguidos por orcas, tubarões e focas esfomeadas, mas o mais estranho nesta história é que a motivação aparente da foca não era a procura de comida (embora tenha devorado parcialmente alguns dos pinguins). Os autores observavam aquela colónia de pinguins desde 1982 e aquele foi o primeiro elefante marinho que viram no meio da colónia. O artigo faz uma descrição de alguns pormenores que não recomendo a espíritos mais sensíveis:Em Outubro de 1997 observámos a foca a atacar um pinguim que fugiu para dentro de uma toca. A foca movimentou a sua cabeça para a entrada da toca e começou a vocalizar. Após mais vocalizações quer do pinguim quer da foca, o pinguim saiu da toca e enfrentou a foca numa postura defensiva.
Um gesto corajoso mas não muito inteligente:Nessa altura a foca elevou-se e largou todo o peso da região inferior do pescoço e da parte anterior do corpo em cima do pinguim. Após se elevar e esmagar o pinguim várias vezes, a foca repousou sobre o pinguim aproximadamente oito minutos e então puxou o pinguim de debaixo do seu corpo e lançou-o no ar. Pouco depois, a foca deslocou-se para uma outra carcaça de pinguim a uns metros de distância. A foca colocou a sua cabeça sobre a carcaça durante alguns minutos e então lançou esta carcaça no ar várias vezes. Aproximadamente 45 minutos mais tarde, a foca voltou ao pinguim que tinha acabado de matar e mais uma vez começou a lançá-lo ao ar, virando-o do avesso, e parecendo comer o pinguim.
Quando em Dezembro de 1997 os autores do artigo percorreram essa área encontraram 27 cabeças e bicos de pinguins. A maioria das carcaças estavam viradas do avesso e os autores só encontravam em geral pele, penas, barbatanas, pés, cabeças e bicos. No ano seguinte a mesma história. Uma foca apareceu na praia a aproximadamente 300 m do local onde tinha sido vista pela primeira vez no ano anterior. Uma semana depois havia já 10 novas carcaças de pinguins, algumas parcialmente comidas. Mais uma vez o mau feitio do pinípede era óbvio:Em Outubro vimos um pinguim dar uma bicada na foca que estava a descansar quase em cima de uma toca onde o pinguim estava a incubar ovos. A foca elevou-se imediatamente e esmagou o pinguim, matando-o. A foca elefante perseguiu pinguins quando eles se aproximavam ou lhe davam bicadas quando tentavam sair do seu local de nidificação.
A foca foi vista desta vez de 17 de Setembro a 1 de Novembro de 1998. Os autores encontraram 49 carcaças de pinguins nas proximidades. A história repetiu-se mais uma vez no ano seguinte, de 19 de Setembro a 5 de Novembro. A carnificina de pinguins continuou:Durante mais de 30 horas de observação, em 1999, vimos a foca a atacar e a matar pinguins, carregando as carcaças ao longo da colónia, virando as carcaças do avesso, e "brincando" com as carcaças, incluindo o lançar das carcaças e o repousar a cabeça em cima delas. Uma vez, a foca perseguiu um pinguim durante aproximadamente 20 metros antes de parar a perseguição.
Desta vez a foca matou pelo menos 12 pinguins. Os autores pensam que se tratou da mesma foca nas três ocasiões, não só pelo comportamento, mas também por apresentar o mesmo tamanho, cor e o mesmo grande desgaste nos caninos inferiores (que se vê claramente na fotografia). Quando a foca deixou de aparecer nos anos seguintes deixaram de aparecer novas carcaças na praia.
Clark e Boersma avançam várias hipóteses para o comportamento da foca. Uma razão poderia ser que a foca via os pinguins como presa. Os autores consideram no entanto pouco provável que esta seja a motivação principal. Estas focas em geral não caçam pinguins e nem todos os pinguins mortos foram comidos pela foca. A chegada da foca coincidiu com a época de acasalamento dos elefantes-marinhos, período em que não se alimentam, e de facto a foca perdia bastante peso durante a estadia em terra. Se o comportamento fosse motivado essencialmente pela fome a foca poderia ter comido muitos mais pinguins.
Outra possibilidade seria a foca ter matado os pinguins para brincar. De facto, parte do comportamento da foca ao lançar pinguins no ar e ao colocar a cabeça sobre eles pode ser encarado sob este prisma. A foca pode também ter reagido a uma fonte de irritação, pois os pinguins pareciam aborrecer a foca, que perseguia ou matava os pinguins que lhe davam bicadas ou de alguma forma a incomodavam. Os autores sugerem ainda que a foca possa ter usado os pinguins como um escape para a agressividade, e um substituto para a frustração, de não ter assegurado um território e um harém de fêmeas durante este período, que corresponde à época de reprodução nesta espécie.
Qualquer que tenha sido a razão a matança deixou as suas marcas. Os autores terminam o artigo com:Em três anos, uma foca matou pelo menos 88 pinguins em Punta Tombo. A foca não voltou em 2000 nem em 2001-2004, sugerindo que terá morrido. Até Março de 2005 a área onde a foca matou os pinguins não tinha voltado à sua densidade anterior, mostrando que um predador, durante um curto intervalo de tempo, pode ter um impacto de longa duração em pinguins em reprodução.
Já agora talvez o elefante-marinho tenha passado por uma experiência com pinguins do tipo desta aqui no Wally & Osborne.
Referências
(ref1) J. Alan Clark e P. Dee Boersma (2006). SOUTHERN ELEPHANT SEAL, MIROUNGA LEONINA, KILLS MAGELLANIC PENGUINS, SPHENISCUS MAGELLANICUS, ON LAND. Marine Mammal Science 22(1): 222-225. Laço DOI.



{kind=link}
{kind=link}