

 Muitos de nós já viram esta imagem: uma grande e gorda lagarta com inúmeros pequenos casulos colados ao corpo. Neste caso trata-se da lagarta da folha do tabaco, de seu nome científico Manduca sexta, e da pequena vespa parasítica Cotesia congregata, que como podem calcular é tida em grande estima pela generalidade das tabaqueiras. Embora imagens de parasitas a sairem dos seus hospedeiros sejam algo que só por si vale a pena partilhar, há muitos outros aspectos interessantes, em particular os resultados de um estudo recente que mostra que a relação entre a planta, a lagarta, e a vespa, é bastante mais complicada do que poderia parecer. As plantas mandam mensagens destinadas às vespas, "dizendo-lhes" que estão a ser atacadas por um tipo específico de lagarta. [... ler mais]
Muitos de nós já viram esta imagem: uma grande e gorda lagarta com inúmeros pequenos casulos colados ao corpo. Neste caso trata-se da lagarta da folha do tabaco, de seu nome científico Manduca sexta, e da pequena vespa parasítica Cotesia congregata, que como podem calcular é tida em grande estima pela generalidade das tabaqueiras. Embora imagens de parasitas a sairem dos seus hospedeiros sejam algo que só por si vale a pena partilhar, há muitos outros aspectos interessantes, em particular os resultados de um estudo recente que mostra que a relação entre a planta, a lagarta, e a vespa, é bastante mais complicada do que poderia parecer. As plantas mandam mensagens destinadas às vespas, "dizendo-lhes" que estão a ser atacadas por um tipo específico de lagarta. [... ler mais]
ntes de falar nisso, no entanto, não resisto a mostrar mais uma imagem que mostra a quantidade enorme de vespas que bem trabucaram outra pobre Manduca. A pesquisa de que vou falar foi efectuada por K. Shiojiri e colegas e publicada na revista PLoS Biology (ref1). A presa neste artigo é a lagarta Mythimna separata, uma peste agrícola que devora vegetais importantes para alimentção humana e para rações animais, como milho, soja e sorgo. O predador é a vespa Cotesia kariyai, uma prima próxima da vespa que mostrei com a larva da folha do tabaco. Eu bem procurei mas não encontrei nenhuma foto com imagens de casulos e pequenas vespas a emergirem desses casulos para esta espécie.
A pesquisa de que vou falar foi efectuada por K. Shiojiri e colegas e publicada na revista PLoS Biology (ref1). A presa neste artigo é a lagarta Mythimna separata, uma peste agrícola que devora vegetais importantes para alimentção humana e para rações animais, como milho, soja e sorgo. O predador é a vespa Cotesia kariyai, uma prima próxima da vespa que mostrei com a larva da folha do tabaco. Eu bem procurei mas não encontrei nenhuma foto com imagens de casulos e pequenas vespas a emergirem desses casulos para esta espécie. Este estudo aborda um aspecto curioso da relação lagarta-vespa. As pequenas vespas são activas durante o dia, enquanto as lagartas em geral só se alimentam durante a noite, escondendo-se durante o dia. O que Shiojiri e colegas testaram foi se é de facto a luminosidade que determina o comportamento das lagartas. É que há outro factor aqui. Quando ameaçadas por herbívoros muitas plantas emitem aquilo que se designa por voláteis das plantas induzidos por herbívoros (VPIH). Este químicos, que se propagam no ar, são em muito casos específicos quanto à espécie que está a atacar a planta. Os VPIHs são vantajosos para as plantas, pois as pequenas vespas parasíticas detectam-nos e procuram a fonte das emissões. A questão que os autores do estudo se puseram foi então a seguinte: as lagartas possuem um comportamento nocturno como resposta ao carácter diurno das vespas, ou respondem às emanações da planta, que apenas são produzidas durante o dia? Este último facto é importante, pois as plantas se atacadas durante a noite não libertam este VPIH específico. Numa tradução livre do resumo do artigo:
Este estudo aborda um aspecto curioso da relação lagarta-vespa. As pequenas vespas são activas durante o dia, enquanto as lagartas em geral só se alimentam durante a noite, escondendo-se durante o dia. O que Shiojiri e colegas testaram foi se é de facto a luminosidade que determina o comportamento das lagartas. É que há outro factor aqui. Quando ameaçadas por herbívoros muitas plantas emitem aquilo que se designa por voláteis das plantas induzidos por herbívoros (VPIH). Este químicos, que se propagam no ar, são em muito casos específicos quanto à espécie que está a atacar a planta. Os VPIHs são vantajosos para as plantas, pois as pequenas vespas parasíticas detectam-nos e procuram a fonte das emissões. A questão que os autores do estudo se puseram foi então a seguinte: as lagartas possuem um comportamento nocturno como resposta ao carácter diurno das vespas, ou respondem às emanações da planta, que apenas são produzidas durante o dia? Este último facto é importante, pois as plantas se atacadas durante a noite não libertam este VPIH específico. Numa tradução livre do resumo do artigo:Embora muitos organismos mostrem ritmos diários no seus padrões de actividade, as causas destes padrões são mal compreendidas. Mostramos aqui que voláteis das plantas afectam o comportamento nocturno da lagarta Mythimna separata. Qualquer que seja o estudo de luminosidade, as lagartas comportavam-se como se estivessem na escuridão se expostas a voláteis emitidas por plantas (infestadas ou não por lagartas congéneres) no escuro. De igual modo,independemente do estado de luminosidade as lagartas comportavam-se se estivessem na luz se expostos a voláteis emitidos por plantas na luz. As lagartas aparentemente utilizam a informação providenciada pelos voláteis das plantas para se informarem sobre o seu ambiente e modularem o seu padrão de actividade diária, dessa forma evitando potencialmente a ameaça do parasitismo.
O método utilizado pelos autores no estudo foi simples, em vez de darem às lagartas plantas vivas, deram-lhes alimentação artificial. As lagartas foram colocadas num pequeno recipiente plástico, com um pequeno abrigo onde se podiam esconder. Os autores verificaram então que as lagartas comportavam-se da mesma forma quer na escuridão quer na claridade, alimentando-se e escondendo-se em números iguais. Tendo estabelecido que a luz não era o factor que controlava o ciclo de actividade das baratas os autores testaram então os efeitos da presença de uma planta de milho. A simples presença da planta, não infestada, levava a que mais 20% das lagartas se escondessem durante o dia, enquanto o número de lagartas escondidas em condições de escuridão caía 30%. O passo seguinte foi utilizar as emanações de plantas infestadas. Ao expor lagartas em condições de claridade ou escuridão a voláteis de plantas atacadas por lagartas (com as plantas em condições de claridade), os autores verificaram que o número de lagartas a esconderem-se aumentava 20 a 40%, quer as lagartas estivessem à luz quer às escuras.
O resultado é fascinante, se a lagarta sente que a planta está a "pedir ajuda" às vespas parte a recolher-se, quer seja noite ou dia. A presença dos VPIH é sentida pela lagarta como indicador imediato de ameaça. O ciclo de vida nocturno é uma consequência indirecta.
Ficha Técnica
Imagens de Manduca sexta, e Cotesia congregata, no topo da página cortesia de R.J. Reynolds Tobacco Company Slide Set, obtidas a partir de www.insectimages.org.
Imagem de Manduca sexta, com inúmeras Cotesia, cortesia de Alton N. Sparks, Jr., The University of Georgia, obtidas através de www.insectimages.org.
Imagem de Cotesia kariyai, cortesia de Dr. James Whitfield e Dr. Michael Sharkey, Hymenoptera institute, Univ. Kentucky.
Imagem de Mythimna separata, cortesia de Liza Gross, PLoS Biology (ref2), ver abaixo.
Referências
(ref1) Shiojiri K, Ozawa R, Takabayashi J (2006) Plant Volatiles, Rather than Light, Determine the Nocturnal Behavior of a Caterpillar. PLoS Biol 4(6): e164. Laço DOI.
(ref2) Gross L (2006) Plants Tell Caterpillars When It's Safe to Forage. PLoS Biol 4(6): e194. Laço DOI.
sexta-feira, maio 19, 2006
Não é da luz que elas têm medo
quarta-feira, maio 17, 2006
A corrida do vampiro

 A contribuição para hoje era sobre astronomia, mas estou com dificuldade em colocar as minhas animações num servidor externo ao blogger. Para não ficar mais um dia em branco resolvi por isso socorrer-me de um artigo antigo (Março 2005) sobre os morcegos vampiros comuns, da espécie Desmodus rotundus. Apesar da má reputação a felpuda criaturinha da imagem até tem um aspecto simpático. O que me leva a falar neste animal não são os sinistros hábitos alimentares, mas sim uma característica algo supreendente: estes morcegos conseguem correr. [... ler mais]
A contribuição para hoje era sobre astronomia, mas estou com dificuldade em colocar as minhas animações num servidor externo ao blogger. Para não ficar mais um dia em branco resolvi por isso socorrer-me de um artigo antigo (Março 2005) sobre os morcegos vampiros comuns, da espécie Desmodus rotundus. Apesar da má reputação a felpuda criaturinha da imagem até tem um aspecto simpático. O que me leva a falar neste animal não são os sinistros hábitos alimentares, mas sim uma característica algo supreendente: estes morcegos conseguem correr. [... ler mais]
Fazem-no com uma passada que embora pouco habitual é bastante eficiente. O artigo foi publicado na Nature (ref1) e é da autoria de Daniel Riskin e John Hermanson, ambos da Universidade de Cornell. Numa tradução livre do resumo:A maioria dos tetrápodes conservaram a locomoção terrestre desde que esta evoluiu na era Paleozóica, mas os morcegos tornaram-se tão especializados para o vôo que perderam practicamente quase toda a habilidade de manobrar em terra. Os morcegos vampiros, que se esgueiram até à sua presa no solo, são uma importante excepção. Mostramos aqui que os morcegos vampiros comuns podem também correr usando uma passada acoplada única, em que os membros anteriores em vez dos posteriores são utilizados para produzir força, uma vez que as asas são muito mais poderosas que as pernas. Esta habilidade para correr parece ter evoluído de forma independente dentro da linhagem dos morcegos.
Os autores colocaram morcegos vampiros numa esteira rolante e verificaram que a baixas velocidades os morcegos eram capazes de caminhar, mas, assim que a velocidade da esteira aumentava, os morcegos desenvolviam algo que só se pode designar por corrida, pois incluía uma fase em que todas as patas estavam fora de contacto com o solo. A passada utilizada era no entanto distinta das formas de corrida utilizadas pelos outros animais de quatro patas, pois eram os braços que produziam a força. As imagens são impressionantes:
As imagens acima não fazem no entanto justiça à fantástica cavalgada do vampiro. A coisa só pode ser apreciada em toda a sua glória neste filme que até é bastante pequeno (569 kbytes). Como mais nenhuma espécie de morcegos mostra esta habilidade os autores concluem que os morcegos vampiros desenvolveram esta forma de corrida de forma independente, isto é, trata-se de uma novidade evolutiva. Os morcegos atingem velocidades razoáveis para um animal do seu tamanho. Na figura o morcego está a correr a puco mais de meio metro por segundo, mas podem atingir cerca de 2 metros por segundo (mais de 7 km/hora).
Agora porque razão terão os morcegos necessidade de correr não é claro. Após a introdução do gado nas Américas pelos humanos no século XVI, os morcegos passaram a alimentar-se preferencialmente nestes animais. Mas os morcegos não são esquisitos, sendo capazes de se alimentarem mesmo de sangue de répteis. É óbvio que correr deve ter alguma utilidade para manobrar em volta das presas, mas sem examinar o comportamento destes vampiros com outros animias, que não o gado, as vantagens do comportamente não são óbvias.
Este animais são muito pequenos, com apenas 8 centímetros de comprimento. Mas há aqui claramente material para pesadelos, não custa imaginar um vampiro gigante a correr desenfreadamente atrás de nós.
Ficha técnica
Imagem do morcego nas paredes da gruta retirada da Wikimedia Commons.
Imagens do morcego a correr e filme retirados da página sobre este estudo na Universidade de Cornell.
Referências
(ref1) Riskin, D.K., and J.W. Hermanson (2005). Biomechanics: Independent evolution of running in vampire bats. Nature 434:292. Laço DOI.
segunda-feira, maio 15, 2006
A montanha de fogo reacende a sua fornalha

 Depois de falar nos hábitos alimentares e no sexo dos anjos, volto hoje a notícias mais terra a terra. Esta é uma imagem do vulcão Gunlung Merapi, que significa montanha de fogo, no centro da ilha de Java, que tem aparecido nas notícias nos últimos dias. A imagem foi tirada pelo instrumento ASTER a bordo do satélite Terra da NASA no dia 26 de Abril de 2006. Podemos ver na imagem uma pluma de vapor a emanar do cimo do vulcão, e rios de lava "seca" de erupções anteriores, com uma cor escura. [...ler mais]
Depois de falar nos hábitos alimentares e no sexo dos anjos, volto hoje a notícias mais terra a terra. Esta é uma imagem do vulcão Gunlung Merapi, que significa montanha de fogo, no centro da ilha de Java, que tem aparecido nas notícias nos últimos dias. A imagem foi tirada pelo instrumento ASTER a bordo do satélite Terra da NASA no dia 26 de Abril de 2006. Podemos ver na imagem uma pluma de vapor a emanar do cimo do vulcão, e rios de lava "seca" de erupções anteriores, com uma cor escura. [...ler mais]
Comprovando que a Indonésia é de facto a terra dos vulcões, uma panorâmica cobrindo mais terreno mostra que este vulcão não stá sozinho, existindo logo ali ao lado um outro vulcão, designado Merbabu.
Trata-se de uma imagem de cor "falsa", em que as regiões com vegetação densa surgem a vermelho e as zonas acinzentadas são regiões em que o terreno foi desbravado, e possivelmente habitadas. A mesma região tinha sido observada em Outubro de 1994, com o radar SIR a bordo do vai-vém espacial Endeavour. Mais uma vez as cores são artificiais. Os dois cumes vulcânicos são evidentes, e as regiões com tons mais verdes e amareladas são fluxo de lava de diferente idades e rugosidades. Seis semanas após estas imagens terem sido tiradas, uma erupção no vulcão Merapi matou 66 pessoas. Uma anterior em 1930 tinha destruído 13 aldeias e tirado a vida a 1,370 habitantes das aldeias redondezas. Não admira que os tremores de terra frequentes no final de Abril e plumas de vapor que mostraram que uma erupção estava iminente tenham levado as autoridades locais a reagir. Afinal o Merapi, localiza-se numa das regiões mais densamente povoadas do mundo, e o número de pessoas a evacuar pode atingir as 80 mil. Segundo os despachos informativos de várias agências alguns milhares de pessoas foram já evacuados. Os fluxos piroclásticos e a possibilidade de torrentes de lama semelhantes aos que acompanharam estas erupções anteriores são neste momento os grandes motivos de preocupação.
Mais uma vez as cores são artificiais. Os dois cumes vulcânicos são evidentes, e as regiões com tons mais verdes e amareladas são fluxo de lava de diferente idades e rugosidades. Seis semanas após estas imagens terem sido tiradas, uma erupção no vulcão Merapi matou 66 pessoas. Uma anterior em 1930 tinha destruído 13 aldeias e tirado a vida a 1,370 habitantes das aldeias redondezas. Não admira que os tremores de terra frequentes no final de Abril e plumas de vapor que mostraram que uma erupção estava iminente tenham levado as autoridades locais a reagir. Afinal o Merapi, localiza-se numa das regiões mais densamente povoadas do mundo, e o número de pessoas a evacuar pode atingir as 80 mil. Segundo os despachos informativos de várias agências alguns milhares de pessoas foram já evacuados. Os fluxos piroclásticos e a possibilidade de torrentes de lama semelhantes aos que acompanharam estas erupções anteriores são neste momento os grandes motivos de preocupação.
Por enquanto a NASA e a UGCS não colocaram imagens à disposição do público. Encontrei contudo esta imagem, obtida também no espaço, que foi tirada por astronautas em 2003.
 O Merapi teve 68 erupções registadas desde 1548, e detém a distinção algo sinistra de ser o vulcão com o maior número de nuvens ardentes: das 68 erupções históricas, 32 tiveram "nuvens ardentes". Uma nuvem ardente é uma grande massa de gases vulcânicos muito quentes que se precipita pelas encostas do vulcão, e que é completamente letal para quem tenha o infortúnio de ser apanhado por ela. Tal como as erupções de Tera, e de Tambora, apresentadas aqui anteriormente, este vulcão está também associado ao colapso de uma sociedade até então próspera. Uma erupção gigantesca em 1006 cobriu toda região central de Java com cinzas vulcânicas. Pensa-se que tessa erupção tenha levado ao colapso da civilização Mataram. Não vou entrar aqui em detalhes, pois o pouco que sei acerca dessa cultura foi através de um documentário televisivo. Dela sobrevivem por exemplo as ruínas de Prambanan, o maior complexo de templos da religião hindu na Indonésia. A imagem da estátua de Ganesh, o deus com cabeça de elefante, é de um dos templos desse santuário. Confesso que gostaria de o visitar, em parte devido ao fascínio que adquiri pela história de Sita. Baixos relevos da história de Sita adornam os muros em redor do santuário, do qual mostro mais uma imagem em baixo.
O Merapi teve 68 erupções registadas desde 1548, e detém a distinção algo sinistra de ser o vulcão com o maior número de nuvens ardentes: das 68 erupções históricas, 32 tiveram "nuvens ardentes". Uma nuvem ardente é uma grande massa de gases vulcânicos muito quentes que se precipita pelas encostas do vulcão, e que é completamente letal para quem tenha o infortúnio de ser apanhado por ela. Tal como as erupções de Tera, e de Tambora, apresentadas aqui anteriormente, este vulcão está também associado ao colapso de uma sociedade até então próspera. Uma erupção gigantesca em 1006 cobriu toda região central de Java com cinzas vulcânicas. Pensa-se que tessa erupção tenha levado ao colapso da civilização Mataram. Não vou entrar aqui em detalhes, pois o pouco que sei acerca dessa cultura foi através de um documentário televisivo. Dela sobrevivem por exemplo as ruínas de Prambanan, o maior complexo de templos da religião hindu na Indonésia. A imagem da estátua de Ganesh, o deus com cabeça de elefante, é de um dos templos desse santuário. Confesso que gostaria de o visitar, em parte devido ao fascínio que adquiri pela história de Sita. Baixos relevos da história de Sita adornam os muros em redor do santuário, do qual mostro mais uma imagem em baixo.
Ficha técnica.
Imagem do ASTER, cortesia do Earth Observatory da NASA, onde pode ser encontrada uma versão de alta resolução.
Imagem do SIR, cortesia do PhotoJournal da NASA, com acesso a versão de alta resolução.
A foto do vulcão tirada do vai-vém espacial é também cortesia do Earth Observatory da NASA.
As imagens de Prambanan, foram retiradas da Wikipédia versão inglesa.
sábado, maio 13, 2006
O alimentar dos anjos


 A última coisa que nos ocorre quando vemos esta cabeça encimada por tentáculos que parecem cornos é que pertence a uma criatura designada por anjo do mar. O porquê do nome fica uma incógnita ainda maior quando se vêem mais tentáculos a sairem da bocarra aberta. Mas é essa mesmo a designação deste ser. Trata-se de mais um exemplar dos fabulosos caracóis voadores de que falei aqui há pouco tempo, neste caso da espécie Clione limacina. A semelhança com um anjo é contudo óbvia quando se vê o bicharoco a nadar. [... ler mais]
A última coisa que nos ocorre quando vemos esta cabeça encimada por tentáculos que parecem cornos é que pertence a uma criatura designada por anjo do mar. O porquê do nome fica uma incógnita ainda maior quando se vêem mais tentáculos a sairem da bocarra aberta. Mas é essa mesmo a designação deste ser. Trata-se de mais um exemplar dos fabulosos caracóis voadores de que falei aqui há pouco tempo, neste caso da espécie Clione limacina. A semelhança com um anjo é contudo óbvia quando se vê o bicharoco a nadar. [... ler mais]
Após uma curta pesquisa pela Internet encontrei dois filmes que mostram o vigoroso e ritmado bater de asas da Clione. Os filmes, que eu recomendo, e até nem são tão grandes como isso, podem ser encontrados aqui (1.7 Mbytes), e aqui (0.5 Mbytes).
Mas não foi a graciosidade com que a criatura se movimenta que me levou a falar nela. O que a leva a ser a criatura do dia foi o facto de eu ter encontrado um artigo de 1992, da autoria de Hermans e Satterlie, no Biol. Bull. (ref1), em que estes autores descrevem a forma como este anjo se alimenta. Os detalhes eram bons demais para não os partilhar. Eu já tinha referido aqui que os anjos se alimentavam de borboletas, e no caso da Clione limacina, a escolha recai sobre a espécie Limacina helicina. Trata-se da única borboleta do mar que vive na região do ártico, onde atinge densidades algo elevadas e é um elemento importante da cadeia alimentar para muitas espécies. A Limacina helicina alimenta-se estendendo uma rede de muco que usa para apanhar algas microscópicas e também alguns pequenos animais. Embora sejam colectivamente designados por pterópodes, as borboletas e os anjos do mar não são parentes tão próximos como isso, formando duas categorias distintas dentro dos gastrópodes. As borboletas pertencem a um grupo cuja designação científica é Thecosomata, os anjos a um grupo designado Gymnosomata.
Eu já tinha referido aqui que os anjos se alimentavam de borboletas, e no caso da Clione limacina, a escolha recai sobre a espécie Limacina helicina. Trata-se da única borboleta do mar que vive na região do ártico, onde atinge densidades algo elevadas e é um elemento importante da cadeia alimentar para muitas espécies. A Limacina helicina alimenta-se estendendo uma rede de muco que usa para apanhar algas microscópicas e também alguns pequenos animais. Embora sejam colectivamente designados por pterópodes, as borboletas e os anjos do mar não são parentes tão próximos como isso, formando duas categorias distintas dentro dos gastrópodes. As borboletas pertencem a um grupo cuja designação científica é Thecosomata, os anjos a um grupo designado Gymnosomata.
Voltemos então ao artigo de Hermans e Satterlie, que no início descrevem em detalhe o elaborado e longo processo para a captura e consumo das presas da Clione. Numa tradução livre do original em inglês: Para a aquisição da presa, a Clione procede rapidamente à extrosão de três pares de apêndices orais, chamados cones bucais, que cercam e aderem à presa. Cada cone bucal tem forma cónica quando retraído mas torna-se mais cilíndrico aquando da extrosão. A extrosão dos cones bucais é devida essencialmente a enchimento hidráulico. A fase de aquisição da presa é seguida por uma fase manipulativa, durante a qual a presa é virada por forma a que a abertura da concha fique sobre a boca da Clione. A manipulação é efectuada pelos cones bucais e é seguida pela fase de consumo, durante a qual a presa é extraída da concha. A extração envolve o uso de dois sacos de ganchos quitinosos especializados que são parte do aparelho bucal. Cada saco de ganchos contêm tufos de ganchos quitinosos recurvados, que são colocados no interior da abertura da concha para agarrar e puxar a presa da sua concha. Os tecidos moles da presa são desalojados por movimentos alternados de protacção e retracção dos sacos de ganchos. O engolir é auxiliado por movimentos de protacção e retracção da rádula, que é também parte do aparelho bucal. Os tecidos moles da presa são engolidos inteiros.
Os tais cones bucais são as coisa que se veêm a sair da boca da Clione na imagem logo no início desta contribuição. Tudo demora cerca de 30 minutos, que são passados essencialmente no manobrar da presa e sobretudo no lento puxar da relutante Limacina da sua concha. O artigo de Hermans e Satterlie é essencialmente dedicado à fase de captura. Se após segurar a presa, a Clione demora bastante tempo até engoli-la, a fase inicial é extraordinariamente rápida.A fase de aquisição da refeição envolve abertura rápida da boca e a extrosão de três pares de cones bucais. A abertura da boca ocorre em 10 a 20 milissegundos, enquanto o enchimento hidrostático dos cones bucais demora 50 a 70 milissegundos.
O processo está ilustrado nas imagens abaixo que os autores obtiveram através da filmagem com uma câmara de alta velocidade, com uma imagem a cada 10 milissegundos. A segunda imagem neste sequência corresponde ao instante do contacto do anjo com a sua presa, uma pobre borboleta segura por uma pinça.
Na terceira imagem, 10 milissegundos após o contacto, o anjo está já a estender os cones bucais, que na quarta imagem já estão estendidos quase ao máximo e já seguram a infeliz presa. É impressionante mas este animal necessita de menos de um décimo de segundo para segurar a vítima após fazer contacto. A Clione não deixa nada ao acaso:Os cones bucais rodeiam a presa e libertam um material viscoso que pode ser utilizado como um suporte adesivo à concha da presa.
A pobre borboleta do mar não tem nenhuma hipótese de fuga. Enfim, o estar segura pela pinça também não ajudou em nada neste caso. O seu destino é ser engolida duma só vez.
Há mais alguns aspectos peculiares destes animais que eu aproveito para partilhar aqui. Uma das características mais curiosas dos Thecosomata como a Limacina helicina é que, embora tenham reprodução sexuada com sexos diferenciados, o sexo do animal depende da idade. As Limacina do ártico começam a vida adulta como machos, mas quando ultrapassam os 5 mm de comprimento mudam de sexo e tornam-se fêmeas. Quanto à velha questão do sexo dos anjos, pois bem os Gymnosomata, como a Clione, são hermafroditas simultâneos, isto é, os dois sexos estão presentes ao mesmo tempo em cada indivíduo. Todos os anjos do mar possuem uma abertura genital feminina e um orgão masculino para a cópula. A coisa mais parecida com pornografia dos anjos que encontrei foi esta foto aqui, que mostra que o acasalamento da Clione é feito com os indivíduos em posição ventral. Infelizmente não a posso colocar aqui porque o autor é muito claro quanto aos direitos.
Esta é possivelmente a minha última incursão no domínio dos caracóis voadores nos tempos mais próximos. Trata-se contudo de criaturinhas fascinantes, e se aparecerem artigos recentes com detalhes interessantes voltarei a elas.
Referências
(ref1) COLIN 0. HERMANS' AND RICHARD A. SATTERLIE (1992). Feeding Behavior in a Pteropod Mollusk, Clione limacina Phipps. Biol. Bull. 182: l-7. Resumo e PDF
sexta-feira, maio 12, 2006
Cefalópodes bebés, pequeninos e amorosos, lançados às feras

 De volta ao Censo da Vida Marinha. Para lá de anjos, borboletas, e elefantes adultos, a equipa de cientistas capturou alguns exemplares de bebés com tentáculos. Os pequenos cefalópodes são forçados a desenvencilharem-se sozinhos no vasto mar dos sargaços, sem auxílio dos seus progenitores. Apesar dos grandes olhos e aspecto amoroso estas criaturas não são propriamente meigas. O título desta contribuição só em parte está correcto. Estes pequenitos podem ter sido lançados às feras, mas para muitos habitantes do zooplâncton as feras são eles. [... ler mais]
De volta ao Censo da Vida Marinha. Para lá de anjos, borboletas, e elefantes adultos, a equipa de cientistas capturou alguns exemplares de bebés com tentáculos. Os pequenos cefalópodes são forçados a desenvencilharem-se sozinhos no vasto mar dos sargaços, sem auxílio dos seus progenitores. Apesar dos grandes olhos e aspecto amoroso estas criaturas não são propriamente meigas. O título desta contribuição só em parte está correcto. Estes pequenitos podem ter sido lançados às feras, mas para muitos habitantes do zooplâncton as feras são eles. [... ler mais]
Os polvos e lulas bebés são predadores activos que buscam e perseguem as suas presas. Para ilustrar esse ponto procurei e encontrei nos arquivos da NOAA imagens que não foram retiradas deste estudo, mas que estão de certa forma relacionadas, pois mostram um parente próximo do par acima, um outro cefalópode bebé (polvo?), a atacar uma borboleta do mar. Ou pelo menos assim parece. Infelizmente as imagens não tinham descrição e fiquei sem saber se a história teve um final feliz para o pequeno cefalópode e infeliz para o gastrópode, ou vice-versa.
Ou pelo menos assim parece. Infelizmente as imagens não tinham descrição e fiquei sem saber se a história teve um final feliz para o pequeno cefalópode e infeliz para o gastrópode, ou vice-versa.
Já que falei de polvos, não resisto a colocar aqui um laço para um filme que mostra um polvo que se assemelha ao Dumbo (o famoso elefante dos desenhos animados). O filme foi obtido a 3 km de profundidade e pode ver-se aqui, cortesia do Censo da Vida Marinha e de Michael Vecchione/NOAA.
quinta-feira, maio 11, 2006
No mar os elefantes gostam de caçar borboletas

 Embora eu considere que uma criatura não precisa de ser bela para se poder considerar fascinante, que qualificativos utilizar quando se encontra uma criatura que parece saída de um pesadelo, como este Oxygyrus keraudreni? Esta, digamos não muito bela, criatura, pertence a um grupo de caracóis marinhos minúsculos (menos de 1cm) que fazem parte do zooplâncton. [... ler mais]
Embora eu considere que uma criatura não precisa de ser bela para se poder considerar fascinante, que qualificativos utilizar quando se encontra uma criatura que parece saída de um pesadelo, como este Oxygyrus keraudreni? Esta, digamos não muito bela, criatura, pertence a um grupo de caracóis marinhos minúsculos (menos de 1cm) que fazem parte do zooplâncton. [... ler mais]
A imagem foi retirada do Censo da Vida Marinha, de que falei aqui ontem. Trata-se de um membro de um grupo de animais chamados heterópodes, também designados por elefantes do mar, devido à espécie de tromba que apresentam. Como se pode adivinhar pelo título desta contribuição, são predadores, gostando em particular das borboletas do mar, de que falei aqui ontem. Estas criaturas apresentam algumas diferenças relativamente a esses caracóis alados, os pterópodes. A concha que apresentam é transparente e em muitos casos formada por uma espécie de cartilagem (conchiolina) e não por aragonite como nas borboletas do mar. Por outro lado o pé não deu lugar a asas mas a uma espécie de remo, cujo abanar impele o animal através da água. Este grande plano de outra espécie destas criaturas, a Atlanta peroni, mostra alguns detalhes dos elefantes do mar. Os olhos são utilizados pelo animal para detectar as suas presas, embora a forma como formam as imagens seja curiosa. Veêm apenas uma tira de cada vez e têm que executar uma série de varrimentos com os olhos para construirem a imagem. Depois de detectarem a sua infeliz presa, que pode ser uma borboleta do mar, utilizam a ventosa e os tentáculos para se fixarem a ela e a manobrarem. A criatura é comida através da "tromba", ou usando uma terminologia mais adequada, a probóscide. Eis em baixo duas fotos de corpo inteiro destes animais. Note-se que os elefantes do mar nadam "às avessas", isto é, com a concha para baixo e o pé-barbatana para cima.
Este grande plano de outra espécie destas criaturas, a Atlanta peroni, mostra alguns detalhes dos elefantes do mar. Os olhos são utilizados pelo animal para detectar as suas presas, embora a forma como formam as imagens seja curiosa. Veêm apenas uma tira de cada vez e têm que executar uma série de varrimentos com os olhos para construirem a imagem. Depois de detectarem a sua infeliz presa, que pode ser uma borboleta do mar, utilizam a ventosa e os tentáculos para se fixarem a ela e a manobrarem. A criatura é comida através da "tromba", ou usando uma terminologia mais adequada, a probóscide. Eis em baixo duas fotos de corpo inteiro destes animais. Note-se que os elefantes do mar nadam "às avessas", isto é, com a concha para baixo e o pé-barbatana para cima.
No mundo destas minúsculas criaturas, onde elefantes e anjos gostam de perseguir e devorar borboletas, há muitos outros animais fascinantes e estranhos. Na verdade o censo da vida marinha é tão interessante que provavelmente vou passar o resto da semana a falar nele.
Há alguns locais na internet dedicados a estes animais. Um deles mostra o aspecto da "boca", a rádula, na extremidade da probóscide, que é arrepiante, tal como se pode ver na figura 3 nesta página.
quarta-feira, maio 10, 2006
Os caracóis voadores, anjos que caçam borboletas
 O Censo da Vida Marinha é uma rede de investigadores de 70 países que estudam a diversidade, distribuição e abundância da vida marinha. Os últimos resultados desse estudo acabam de ser publicitados (pdf), incluindo a sequenciação genética de 220 espécies de zooplâncton encontradas entre 1 e 5 km de profundidade. As fotos destas criaturas, muitas delas com dimensões de alguns milímetros ou inferiores, são espectaculares. Um dos grupos que me chamou mais a atenção foram espécies de caracóis nadadores, um grupo designado por pterópodes. Exacto, ptero como em algo que tem asas. [... ler mais]
O Censo da Vida Marinha é uma rede de investigadores de 70 países que estudam a diversidade, distribuição e abundância da vida marinha. Os últimos resultados desse estudo acabam de ser publicitados (pdf), incluindo a sequenciação genética de 220 espécies de zooplâncton encontradas entre 1 e 5 km de profundidade. As fotos destas criaturas, muitas delas com dimensões de alguns milímetros ou inferiores, são espectaculares. Um dos grupos que me chamou mais a atenção foram espécies de caracóis nadadores, um grupo designado por pterópodes. Exacto, ptero como em algo que tem asas. [... ler mais]
De facto esses animais fazem jus ao nome pois "voam" através das águas com as suas "asas". Essas "asas", mais correctamente chamadas parapódios, são homólogas aos tradicionais pés dos outros gastrópodes. Podem ver-se em baixo mais algumas espécies destes peculiares caracóis alados.
Na verdade estas fantásticas criaturas formam dois grupos distintos. O primeiro são os Thecosomata, também chamados "borboletas do mar", que possuem concha calcária e são em geral herbívoros, capturando o fitoplâncton com auxílio de uma espécie de rede de muco. Um exemplo é a Cuvierina columnella, que se mostra abaixo em duas poses, uma retraída na sua concha, e noutra mais activa com as suas "asas" de fora. Esta espécie é extremamente comum, incluindo nos mares do Brasil, como descobri num artigo na Revista Brasileira de Zoologia (ref1), de Maria Eduarda de Larrazábal e Valdeni Soares de Oliveira.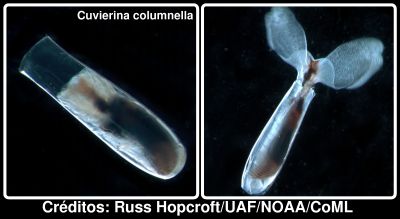
 O outro grupo de pterópodes são os Gymnosomata, os "anjos do mar". O nome engana, estas criaturas são predadores ferozes. Na verdade muitos grupos de Gymnosomata são especialistas em perseguir e capturar as pacíficas borboletas do mar. Os anjos marinhos não possuem concha, apresentam asas semelhantes aos outros pterópodes, e um orgão bucal adequado a capturar e devorar as suas infelizes presas, com ganchos, glândulas pegajosas, tentáculos e mesmo ventosas. São também muito fotogénicos, como se pode ver pelo exemplo da Crucibranchaea macrochira que se mostra aqui ao lado.
O outro grupo de pterópodes são os Gymnosomata, os "anjos do mar". O nome engana, estas criaturas são predadores ferozes. Na verdade muitos grupos de Gymnosomata são especialistas em perseguir e capturar as pacíficas borboletas do mar. Os anjos marinhos não possuem concha, apresentam asas semelhantes aos outros pterópodes, e um orgão bucal adequado a capturar e devorar as suas infelizes presas, com ganchos, glândulas pegajosas, tentáculos e mesmo ventosas. São também muito fotogénicos, como se pode ver pelo exemplo da Crucibranchaea macrochira que se mostra aqui ao lado.
Referências
(ref1) Maria Eduarda de Larrazábal; Valdeni Soares de Oliveira (2003). Thecosomata e Gymnosomata (Mollusca, Gastropoda) da cadeia Fernando de Noronha, Brasil. Rev. Bras. Zool. vol.20 no.2. Laço HTML.
segunda-feira, maio 08, 2006
É sempre difícil mudar de casa

 Chega uma altura na vida de um enxame de abelhas domésticas, Apis mellifera, em que o grupo tem que encontrar um novo local onde viver, construir uns favos, e criar uns quantos milhares de larvas. À raínha convém preservar as forças para pôr ovos, e enviar todos os membros do enxame numa busca de habitações seria um desperdício da energia da colónia. A solução mais óbvia é enviar umas quantas obreiras como batedores, e depois escolher entre os locais que elas encontrem, minimizando o tempo de busca e os recursos dispendidos pela colónia. Neste processo as abelhas beneficiam da peculiar forma de comunicar informações entre os membros do grupo, a famosa dança das abelhas, que as obreiras utilizam para informar os outros membros da colónia acerca da localização de campos de flores. [... ler mais]
Chega uma altura na vida de um enxame de abelhas domésticas, Apis mellifera, em que o grupo tem que encontrar um novo local onde viver, construir uns favos, e criar uns quantos milhares de larvas. À raínha convém preservar as forças para pôr ovos, e enviar todos os membros do enxame numa busca de habitações seria um desperdício da energia da colónia. A solução mais óbvia é enviar umas quantas obreiras como batedores, e depois escolher entre os locais que elas encontrem, minimizando o tempo de busca e os recursos dispendidos pela colónia. Neste processo as abelhas beneficiam da peculiar forma de comunicar informações entre os membros do grupo, a famosa dança das abelhas, que as obreiras utilizam para informar os outros membros da colónia acerca da localização de campos de flores. [... ler mais]
Esta dança é também importante na escolha da habitação. O processo de tomada de decisão é descrito num artigo no American Scientist, da autoria de Thomas D. Seeley e colegas. Numa tradução livre do resumo:Um dos pontos de viragem na vida de uma abelha melífera é quando a abelha-raínha abdica da sua colmeia a favor da sua raínha filha, e leva consigo metade das obreiras partindo para começar um novo ninho. O processo de escolha de uma nova casa pelas abelhas que partem parece levar algum tempo. Com uma pesquisa cuidadosa, Thomas Seeley e colegas revelaram a forma como um enxame chega a uma decisão. Não se trata exactamente de uma democracia, mas sim de uma questão de atingir um limiar de abelhas que favorecem um determinado local utilizando a sua dança. Num grupo de 10,000 abelhas, algumas centenas buscam locais para ninho, mas bastam apenas 10 a 20 abelhas num local para que o enxame se movimente nessa direcção. Através de experimentação e modelação matemática, o grupo de Seeley mostrou que o método das abelhas é eficiente a pesar entre a necessidade de escolher uma casa rapidamente e entre escolher o ninho ideal.
O processo de escolha pode ser descrito em três passos, que funcionam desde que a abelha batedora seja "honesta" quanto ao que achou. A qualidade do abrigo é definida em função da abertura da entrada, que não deve ser demasiado estreita, nem demasiado larga, e do volume que deve ser capaz de acolher eficientemente o enxame e futura colmeia. Quando encontra um local adequado para um ninho, a batedora volta ao enxame e inicia uma dança, executando uma série de oitos, que indica a direcção até ao ninho potencial. O factor determinante para o processo de escolha é que as abelhas ajustam a intensidade da dança em razão da qualidade do abrigo que encontraram. Quanto melhor o abrigo, mais vigorosa é a dança e mais tempo dura. Ora outras abelhas vão inspecionar estes locais e quando voltam executam a sua própria dança, que reflecte o seu próprio "julgamento" quanto ao local. Isto evita erros de apreciação individuais e, como o número de novas recrutas que vão ver um determinado local depende da duração e vigor da dança, faz com que os locais indicados como bons sejam progressivamente visitados por uma maior fracção de abelhas. O resultado é que ao fim de algumas horas os locais menos bons são efectivamente excluídos da "competição" e o enxame parte na direcção de um local que em geral não foi visitado por mais de 20 abelhas.
Esta forma de escolha é rápida e eficiente e tende a produzir bons resultados. As abelhas que voltam estão no fundo a formar correntes de opinião baseadas no seu "julgamento" de quão bom é o abrigo. É claro que estamos a falar de animais com um número reduzido de neurónios e os conceitos de escolha e decisão não são de todo equivalentes aos humanos. O que o estudo mostra é que em sistemas complexos, com um grande número de indivíduos, o caminho para tomar uma escolha acertada nem sempre passa pela consulta a todos os indivíduos. O sistema é apesar de tudo "democrático" num certo sentido, pois os indivíduos que levam o enxame a optar por um rumo não formam nenhum grupo com interesses particulares, mas são obreiras como as outras, e o enxame segue estas abelhas sem ser coagido. É no entanto muito diferente do critério "democrático" adoptado pelas baratas, que vivem em pequenas comunidades, e de que falámos aqui há algum tempo. Em relação à forma incipiente de gregarismo das baratas, que vivem em grupos de poucas dezenas de indivíduos, as abelhas são bastante diferentes. Trata-se de insectos eussociais que vivem em colónias de milhares de indivíduos, e o critério de escolha visa economizar os recursos da colónia. O algoritmo que dita este comportamento é muito simples e ao alcance de animais com um poder mental relativamente reduzido, e é isso que o torna tão interessante.
Uma vez estabelecidas no novo local, as abelhas podem começar a tratar de outros aspectos do futuro da nova colónia. A imagem abaixo, mostra um grande plano de uma destas criaturas, um dos poucos insectos apreciados pelos seres humanos.
Aquilo que eu considero particularmente fascinante neste tipo de estudos são os aspectos relativos à dança das abelhas. Há claramente a transmissão de informação, mas o comportamento vai para além disso, em particular há transmissão de um "juízo" que visa influenciar o comportamento futuro do grupo. Não resisto por isso a indicar uma contribuição no blog de John Hawks, que cita um artigo de Lestel de 2002 (ref2), que por sua vez discute um artigo de Haldel de 1953, sobre o significado da dança das abelhas na busca de alimento. Numa tradução livre de uma parte que eu considero particularmente interessante:Na colmeia, as abelhas fazem indubitavelmente movimentos que originam respostas noutras abelhas; mas as primeiras não estão necessariamente a comunicar informação acerca da nova fonte de alimento. Alguns desses movimentos podem ser encarados como formas de transmitir a próxima acção. Haldane chegou à conclusão lógica que a distinção entre comunicação e acção não é tão clara como tinha parecido. Não só podem os animais transmitir movimentos indicando intenção, como podem também responder a eles. Quanto mais ritualizados os movimentos, mais fácil é responder-lhes. Haldane sugeriu então que mais que a comunicação de uma mensagem, a dança das abelhas era um movimento de intenções altamente ritualizado que se realizava antes de abandonar a colmeia e que levava outras abelhas a partir da mesma forma. A dança da abelha melífera pode assim ser interpretada como a previsão dos seus movimentos futuros em vez da descrição dos seus movimentos passados. Haldane considerava que as danças das abelhas eram interessantes devido à sua "ambiguidade temporal", que as torna simultaneamente profecias e histórias (Lestel 2002:45-46).
"Simultaneamente profecias e histórias" é uma expressão particularmente feliz.
Referências
(ref1) Thomas D. Seeley, P. Kirk Visscher, Kevin M. Passino (2006). Group Decision Making in Honey Bee Swarms. When 10,000 bees go house hunting, how do they cooperatively choose their new nesting site? American Scientist, Volume 94, Number 3, Page 220. Laço DOI
(ref2) Lestel D. (2002). The biosemiotics and phylogenesis of culture. Soc Sci Information 41:35-68. Resumo
domingo, maio 07, 2006
O canto do cardeal visto aos raios-X
 A criatura de hoje é uma notícia um bocado antiga, de um artigo saído em Abril. Contudo, ver um filme em raios-X de uma ave a cantar, é algo que eu tinha que partilhar com os dois ou três leitores que vêm aqui regularmente. O passaroco em questão é o cardeal da Virgínia, de seu nome científico Cardinalis cardinalis, uma simpática ave canora com um belíssimo casaco de penas. O estudo científico em questão, que foi publicado nos Proceedings da National Academy of Sciences (ref1), por Tobias Riede e colegas, tinha como objectivo perceber de que forma os pássaros modulam o seu canto. [... ler mais]
A criatura de hoje é uma notícia um bocado antiga, de um artigo saído em Abril. Contudo, ver um filme em raios-X de uma ave a cantar, é algo que eu tinha que partilhar com os dois ou três leitores que vêm aqui regularmente. O passaroco em questão é o cardeal da Virgínia, de seu nome científico Cardinalis cardinalis, uma simpática ave canora com um belíssimo casaco de penas. O estudo científico em questão, que foi publicado nos Proceedings da National Academy of Sciences (ref1), por Tobias Riede e colegas, tinha como objectivo perceber de que forma os pássaros modulam o seu canto. [... ler mais]
Numa tradução livre do resumo:No discurso humano, o som é gerado pela laringe e modificado por movimentos do tracto vocal superior, que actua como um filtro ressonante concentrando a energia na proximidade de frequências específicas, ou formandos, essenciais no reconhecimento da fala. Apesar da sua importância potencial na comunicação vocal, pouco se conhece acerca da presença de filtros ajustáveis nos tractos vocais de outros vertebrados. A qualidade tonal de muitos dos cantos das aves, nos quais os harmónicos superiores têm relativamente pouca energia, depende da filtragem da fonte vocal, mas a natureza do filtro é controversa. As hipóteses correntes tratam o tracto vocal das aves canoras como um tubo rígido com uma ressonância que é modulada pela correcção na extremidade devida a uma abertura variável do bico.
É sempre curioso ver o tipo de coisas aparentemente simples que desconhecemos. O modelo que se admitia para os pássaros era no fundo do tipo de uma flauta, um tubo rígido em que tapando os orifícios com os dedos se muda a tonalidade da música. Nos pássaros algo semelhante seria conseguido abrindo mais ou menos o bico. Mas o mais interessante de tudo são as técnicas que os cientistas usam para testar estas hipóteses.Através de cinematografia de raios-X de aves canoras, mostramos que o canto da ave é acompanhado por movimentos cíclicos do esqueleto hióide e por mudanças no diâmetro da extremidade craniana do esófago que mantêm um relação inversa entre o volume da cavidade orofaríngica e esófago e a frequência fundamental do canto. Um modelo acústico computacional indica que este padrão motor relacionado com o canto ajusta a ressonância da cavidade orafaríngica-esofágica para acompanhar de forma activa a frequencia fundamental do canto.
Por estranho que possa parecer, os pássaros cantam de uma forma muito semelhante à nossa, ajustando o volume da "garganta". O resultado científico é interessante mas tenho que confessar que o que me fascinou mesmo foi ver um filme da radiografia do animal a cantar. Eis aqui algumas imagens que eu "roubei" da animação em questão, que mostram a transição de um cardeal "vestido" para uma criatura de estranho aspecto. A última imagem representa uma visão de frente.
Mas o melhor mesmo é ver a animação, que até nem é tão grande como isso (~10 Mbytes) que pode ser obtida no laço DOI dado com a referência. Uma versão reduzida pode ser vista aqui, juntamente com um pequeno comunicado de imprensa (em inglês). É claro que para os leitores mais antigos do Cais de Gaia, a questão que se põe é: e então o canto das ratazanas?
Referências
(ref1) Tobias Riede, Roderick A. Suthers, Neville H. Fletcher, William E. Blevins (2006). Songbirds tune their vocal tract to the fundamental frequency of their song, PNAS, vol. 103, no. 14, 5543-5548. Laço doi
quinta-feira, maio 04, 2006
A oliveira e a idade do Minotauro
 Este bocado de madeira, com cerca de um metro de comprimento, pode não parecer nada de especial, mas permitiu fixar com razoável exactidão uma das grandes erupções vulcânicas da antiguidade. Já discuti, nalgumas contribuições anteriores, ocasiões em que a arqueologia encontra a vulcanologia, em geral mostrando os efeitos desastrosos para as populações da área afectada pelo cataclismo. Tais foram os casos da erupção do Vesúvio na região de Avelino, e de Tambora na ilha de Sumbawa. A erupção de que vamos falar hoje foi também devastadora e aconteceu em pleno mediterrâneo. Comecemos por recuar 140 anos, até 1866. Nesse ano, Robert Zahn, ao escavar uma camada de pedra-pomes na ilha de Tera (Santorini), descobriu uma "pompeia da Idade do Bronze". Escavações posteriores, em especial em 1967 num local chamado Akrotiri, pelo Prof. Spyridon Marinatos, trouxeram à luz do dia vestígios de uma grande cidade, com edifícios de vários andares, ruas e praças. Estes eram traços de uma cultura sofisticada e requintada, com ligações à cultura minóica então existente em Creta. O "então" significa há mais de 3500 anos.[... ler mais]
Este bocado de madeira, com cerca de um metro de comprimento, pode não parecer nada de especial, mas permitiu fixar com razoável exactidão uma das grandes erupções vulcânicas da antiguidade. Já discuti, nalgumas contribuições anteriores, ocasiões em que a arqueologia encontra a vulcanologia, em geral mostrando os efeitos desastrosos para as populações da área afectada pelo cataclismo. Tais foram os casos da erupção do Vesúvio na região de Avelino, e de Tambora na ilha de Sumbawa. A erupção de que vamos falar hoje foi também devastadora e aconteceu em pleno mediterrâneo. Comecemos por recuar 140 anos, até 1866. Nesse ano, Robert Zahn, ao escavar uma camada de pedra-pomes na ilha de Tera (Santorini), descobriu uma "pompeia da Idade do Bronze". Escavações posteriores, em especial em 1967 num local chamado Akrotiri, pelo Prof. Spyridon Marinatos, trouxeram à luz do dia vestígios de uma grande cidade, com edifícios de vários andares, ruas e praças. Estes eram traços de uma cultura sofisticada e requintada, com ligações à cultura minóica então existente em Creta. O "então" significa há mais de 3500 anos.[... ler mais]
A atestar o "saber-viver" da população da ilha, mesmo nessa data tão recuada já tinham água canalizada, que incluía mesmo canos de água quente, provavelmente de origem geo-térmica. As casas estavam decoradas com belíssimas pinturas, que retratavam cenas da vida de todos os dias, ao contrário do que era habitual para a época, em que os temas usuais eram em geral de inspiração religiosa. Mostro em baixo dois desses frescos, um de crianças a lutar uma forma qualquer de pugilismo, e outro, de um pescador com uma farta captura.
Este tipo de imagens, e muitas outras recuperadas no local, retratos de uma vida próspera e tranquila, dão um carácter ainda mais avassalador à catástrofe que se abateu sobre a ilha. A imagem abaixo, que mostra uma pequena frota de barcos, rodeados de golfinhos, dá-me a sensação algo estranha de que a Santorini da Idade do Bronze seria um local que eu gostaria de ter visitado. Não sei porquê mas parece-me uma sociedade feliz.
 Santorini é uma caldeira vulcânica rodeada de um conjunto de pequenas ilhas, a maior das quais se chama Tera. Antes da erupção que soterrou a cidade de Akrotiri, a caldeira era rodeada por uma ilha circular com apenas uma pequena entrada. A erupção do vulcão foi devastadora para as populações locais, com uma coluna de cinzas e gases vulcânicos a elevar-se a cerca de 40 km, e enterrou a cidade debaixo de 60 metros de materiais vulcânicos. Vestigíos dessa erupção foram encontrados em locais como a China, Gronelândia e a costa oeste da América do Norte. A erupção foi acompanhada de Tsunamis com mais de 10 metros de altura que atingiram a ilha de Creta a mais de 100 km dali, e que podem ter precipitado o colapso da civilização minóica. Tendo sido sentida em todo o mediterrâneo oriental, a erupção é um acontecimento importante para comparar as cronologias aceites pelos historiadores para as várias culturas da região. O problema é que os vários métodos para determinar a idade da erupção dão resultados contraditórios.
Santorini é uma caldeira vulcânica rodeada de um conjunto de pequenas ilhas, a maior das quais se chama Tera. Antes da erupção que soterrou a cidade de Akrotiri, a caldeira era rodeada por uma ilha circular com apenas uma pequena entrada. A erupção do vulcão foi devastadora para as populações locais, com uma coluna de cinzas e gases vulcânicos a elevar-se a cerca de 40 km, e enterrou a cidade debaixo de 60 metros de materiais vulcânicos. Vestigíos dessa erupção foram encontrados em locais como a China, Gronelândia e a costa oeste da América do Norte. A erupção foi acompanhada de Tsunamis com mais de 10 metros de altura que atingiram a ilha de Creta a mais de 100 km dali, e que podem ter precipitado o colapso da civilização minóica. Tendo sido sentida em todo o mediterrâneo oriental, a erupção é um acontecimento importante para comparar as cronologias aceites pelos historiadores para as várias culturas da região. O problema é que os vários métodos para determinar a idade da erupção dão resultados contraditórios.
Com base, por exemplo, em vestígios de olaria encontrados no local, muito semelhantes ao encontrados no Egipto em regiões que pertenciam ao Império Novo, alguns arqueólogos colocavam a data da erupção próximo de 1500 AC, enquanto a evidência de amostras de gelo, anéis de árvores e datas de medições por radiocarbono indicavam uma data 100 anos anterior. O problema é mais sério do que parece, pois, se as datas mais antigas se mostrarem acertadas, pode ser necessário rever a cronologia egípcia, algo que sempre se pensou estar solidamente estabelecido.
Até recentemente as medições da abundância de carbono-14 eram baseadas em sementes encontradas no local. Ora as datações utlizando as sementes são pouco precisas, pois cada semente dá informações sobre um único ano. Os valores de carbono-14 têm que ser comparados com uma curva, que neste período tem altos e baixos, e os melhores valores indicavam um intervalo de idades possível de 1663 a 1599 AC. Um ponto só não permite uma boa calibração, o que é preciso é algo que forneça uma sequência de pontos. É aqui que entra o fragmento de oliveira encontrado em Santorini. Num artigo na Science (ref1) de Walter Friedrich e colegas esse ramo é utilizado para estabelecer a data da erupção. Numa tradução livre do resumo:Foi possível efectuar uma datação precisa e directa da erupção Minóica en Santorini (Tera) na Grécia, um marco temporal global da Idade do Bronze, através da descoberta de um ramo de oliveira, enterrado na posição que tinha em vida, nas cinzas e pedra-pomes em Santorini.
Este é um facto importante. A árvore estava viva antes da erupção. Os autores baseiam isso no facto de na vizinhança imediata do ramo se encontrarem restos de folhas e azeitonas. O bocado de madeira encontrada incluia a casca da árvore, e os anéis de crescimento imediatamente anteriores. Mesmo numa árvore viva, os anéis interiores, onde já não corre seiva elaborada, são constituídos por células mortas, e os diferentes anéis correspondem a diferentes anos, com diferentes concentrações de carbono-14. Os autores possuíam assim uma sequência relativamente grande para ajustar às flutuações da curva de calibração carbono-14, e não um único ponto como no caso das sementes, e conseguiram uma data bastante exacta para a idade da casca da árvore, e logo para a data em que ocorreu a erupção.Aplicámos o chamado método de ajuste das oscilações a uma sequência de carbono 14 proveniente de segmentos de anéis da árvore para restringir a data da erupção ao intervalo 1627-1600 AC com 95.4% de probabilidade. O nosso resultado está nos limites dos resultados de vários métodos de datação anteriores, menos precisos, e menos directos, mas é um século mais recente que as datas obtidas a partir das cronologias egípcias tradicionais.
É óbvio que a descoberta vai ser alvo de debate, mas é apoiada por um estudo de um outro artigo de que talvez fale aqui na próxima semana.
Referências
(ref1) Walter L. Friedrich, Bernd Kromer, Michael Friedrich, Jan Heinemeier, Tom Pfeiffer, Sahra Talamo (2006). Santorini Eruption Radiocarbon Dated to 1627-1600 B.C. Science, Vol. 312. no. 5773, p. 548. Laço DOI.
terça-feira, maio 02, 2006
Naturalmente resistentes
 Tal como tinha prometido, discuto hoje um descoberta recente acerca do vector da malária, o mosquito Anopheles gambiae. Todos os avanços no conhecimento do vector desta terrível doença são importantes. Aliás, uma das formas de acabar com a malária seria erradicar o vector, mas isso tem-se mostrado difícil. Uma outra opção para acabar com a doença seria substituir o mosquito na natureza por uma estirpe imune à infecção pelo Plasmodium falciparum, e alguns investigadores trabalham exactamente nisso, utilizando engenharia genética. Ora um estudo recente mostra que pelos vistos formas dessas já existem na natureza, e na verdade os mosquitos susceptíveis ao plasmódio serão mesmo uma minoria. [... ler mais]
Tal como tinha prometido, discuto hoje um descoberta recente acerca do vector da malária, o mosquito Anopheles gambiae. Todos os avanços no conhecimento do vector desta terrível doença são importantes. Aliás, uma das formas de acabar com a malária seria erradicar o vector, mas isso tem-se mostrado difícil. Uma outra opção para acabar com a doença seria substituir o mosquito na natureza por uma estirpe imune à infecção pelo Plasmodium falciparum, e alguns investigadores trabalham exactamente nisso, utilizando engenharia genética. Ora um estudo recente mostra que pelos vistos formas dessas já existem na natureza, e na verdade os mosquitos susceptíveis ao plasmódio serão mesmo uma minoria. [... ler mais]
Esta surpreendente descoberta é descrita num artigo na Nature (ref1) de Michelle M. Riehle e colegas. Numa tradução livre do resumo:Pesquisámos uma população de Anopheles gambiae, numa zona de transmissão de malária na África Ocidental, em busca de regiões nos genes capazes de controlar a infecção do mosquito com o parasita da malária, Plasmodium falciparum, que ocorram em condições naturais.
Os autores começaram por isolar fêmeas de mosquito em habitações humanas em áreas rurais no Mali, e utilizaram a prole dessas fêmeas como estirpes para os estudos genéticos. Como cada fêmea do Anopheles gambiae copula uma só vez, cada estirpe de mosquitos utilizada é a prole de um único par, o que é importante para mapear as variantes nos alelos que possam ter efeitos no desenvolvimento do parasita. Essas diferentes estirpes foram então alimentadas com sangue de um único aldeão de uma dessas vilas, aldeão essa que se encontrava infectado com malária.
Ao fim de 7 ou 8 dias os mosquitos que se tinham alimentado do sangue foram dissecados para contar o número de oocistos, uma das fases do desenvolvimento do parasita da malária, que se encontravam no interior de cada mosquito. Isso permitiu aos autores caracterizar cada uma das estirpes quanto à resistência à malária: quantos menos oocistos, mais resistente era a estirpe. Para além disso os autores sequenciaram o ADN de cada um dos mosquitos dissecados, o que permitiu identificar os alelos associados aos diferentes graus de infecção. Das 101 estirpes estudadas pelos autores, 27 foram alvo dos estudos genéticos por terem pelo menos 20 mosquitos, dos quais pelo menos 30% com oocistos. Os autores notam que, curiosamente, outras 20 estirpes não apresentaram qualquer indivíduo com oocistos, embora se tenham alimentado de sangue infectado, o que mostra que a resistência ao plasmódio é comum nos mosquitos. Através da análise genética das 27 estirpes os autores conseguiram então mostrar que a resistência ao plasmódio está localizada numa região relativamente pequena do genoma, num único cromossoma.Os locais de maior resistência ao Plasmodium agrupam-se muma pequena região do cromossoma 2L e cada local explica pelo menos 89% dos mosquitos em estirpes independentes. Em conjunto, esse conjunto de locais formam uma ilha de resistência ao Plasmodium que explica a maior parte da variação genética para a infecção de mosquitos por parasitas da malária na natureza.
Esta segregação, e o elevado nível de resistência encontrado levam os autores a notar nas conclusões:A mais notável característica da resistência observada nos mosquitos é que segrega como um traço mendeliano simples a frequências razoavelmente elevadas em genótipos naturais selecionados de forma aleatória. É interessante notar que muitas estirpes de mosquitos eliminaram completamente o parasita apesar de alimentadas com sangue infectado. Especulamos que o fenótipo-tipo do mosquitos selvagens é a forma resistente, e que a susceptibilidade deve ser atribuída a pontos específicos de falha ou perda de função no sistema imunitário do mosquito.
A existência de um mecanismo genético relativamente simples a controlar a variação natural de susceptibilidade nos mosquitos leva os autores a sugerir que uma forma de controlar a malária seria encontrar um agente que mate preferencialmente os mosquitos infectados com o plasmódio. Fazem então referência aos dois artigos de que falei numa contribuição anterior:Fungos entomopatogénicos podem matar de forma desproporcionada mosquitos infectados com plasmódio quando comparados com não infectados, e podem assim ter as propriedades necessárias a um agente selectivo para transformar as populações do vector numa forma resistente ao Plasmodium. Uma tal estratégia de engenharia evolucionária não exigiria a introdução de nova informação genética em populações naturais do vector.
Ou, dito de outra forma, em vez de tentarmos introduzir uma nova variante do mosquito produzida em laboratório através da engenharia genética, talvez seja mais práctico utilizar os fungos para provocar a extinção das variantes susceptíveis ao plasmódio. Existe muita informação em português na Internet acerca da malária, incluindo algumas páginas de especialistas que descrevem trabalho em curso. Por exemplo, em Portugal estão a decorrer alguns projectos de investigação em malária, no Instituto de Medicina Molecular, sob a direcção de Maria Manuel Mota, em cujas páginas se pode encontrar alguma informação e os laços institucionais de referência.
Referências
(ref1) Michelle M. Riehle, Kyriacos Markianos, Oumou Niaré, Jiannong Xu, Jun Li, Abdoulaye M. Touré, Belco Podiougou, Frederick Oduol, Sory Diawara, Mouctar Diallo, Boubacar Coulibaly, Ahmed Ouatara, Leonid Kruglyak, Sékou F. Traoré, Kenneth D. Vernick (2006). Natural Malaria Infection in Anopheles gambiae Is Regulated by a Single Genomic Control Region. Science 28, Vol. 312. no. 5773, pp. 577-579. Laço DOI.


