

 A Science desta semana tem um artigo interessante sobre dois velhos conhecidos nossos, o Anopheles e o Plasmodium, mas estou com pouco tempo para preparar um blog, pelo que a discussão desse artigo fica para a semana. Tenho contudo material de reserva sobre dois artigos mais antigos sobre esse tema, que introduzem um terceiro ingrediente biológico na equação: fungos, mais exactamente as espécies Beauveria bassiana e Metarhizium anisopliae. Quando um esporo de um destes fungos contacta com um mosquito o resultado é escabroso. O mosquito sofre uma morte lenta, que leva de 10 a 12 dias, sendo coberto por uma camada branca de aspecto peludo. [... ler mais]
A Science desta semana tem um artigo interessante sobre dois velhos conhecidos nossos, o Anopheles e o Plasmodium, mas estou com pouco tempo para preparar um blog, pelo que a discussão desse artigo fica para a semana. Tenho contudo material de reserva sobre dois artigos mais antigos sobre esse tema, que introduzem um terceiro ingrediente biológico na equação: fungos, mais exactamente as espécies Beauveria bassiana e Metarhizium anisopliae. Quando um esporo de um destes fungos contacta com um mosquito o resultado é escabroso. O mosquito sofre uma morte lenta, que leva de 10 a 12 dias, sendo coberto por uma camada branca de aspecto peludo. [... ler mais]
O esforço de erradicação da malária tem passado em grande parte pela utilização de pesticidas químicos, que embora eficazes tem vindo a ter problemas pois os mosquitos têm desenvolvido resistência aos pesticidas utilizados. Métodos biológicos têm sido experimentados com algum sucesso para as larvas destes mosquitos, mas até recentemente não se tinha tentado implementar um controle biológico de mosquitos adultos. É aí que entram os fungos em questão. O primeiro artigo da Science de Junho de 2005 (ref1), é da autoria de Simon Blanford e colegas. Numa tradução livre do resumo:Usando um modelo da malária de roedores, verificámos que a exposição a superfícies tratadas com fungos entomopatogénicos a seguir a uma refeição com sangue infectado reduzia o número de mosquitos capazes de transmitir malária por um factor de cerca de 80. A infecção pelo fungo, conseguida através de contacto com superfícies sólidas ou redes em durações dentro dos períodos típicos de descanso após alimentação, foi suficiente para causar mais de 90% de mortalidade. As taxas de mortalidade diárias aumentaram dramaticamente por volta do tempo de maturação do esporozoíto, e os mosquitos infectados mostraram propensão reduzida para sugar sangue. Aspersões residuais de biopesticidas fungais poderão substituir ou complementar pesticidas químicos para o controle da malária, em particular nas áreas com elevada resistência ao insecticida.
Infectar estes mosquitos é relativamente fácil, pois como todos sabemos, depois de se empaturreram com o nosso sangue os mosquitos têm a mania de pousarem nas paredes. Um dos resultados são as características manchas de sangue nas paredes quando nos decidimos vingar e esborrachá-los. Daí que baste pulverizar as superfícies com esporos destes fungos, mesmo em doses muito baixas. Eu coloquei o ênfase em dois benefícios extra que os autores do estudo notaram. O parasita da malária, o Plasmodium, tinha o seu desenvolvimento perturbado nos mosquitos infectados pelo fungo. Além disso os mosquitos infectados pelo fungo mostravam menor propensão a alimentarem-se, o que torna menos provável que adquiram o plasmódio. São estes aspectos que leva à redução por um factor de 80 na taxa de infecção. Embora o intervalo de 10 a 12 dias até à morte dos mosquitos pareça longo, é adequado para combater a malária. Isto porque o parasita da malária demora cerca de 14 dias a desenvolver-se da fase de gametonte no sangue que o mosquito ingeriu, até à fase infecciosa para os seres humanos, o esporozoíto, que vive nas glândulas salivares do mosquito.
Neste primeiro artigo foi estudado o mosquito Anopheles stephensi, que é o vector envolvido na transmissão da malária dos ratos, causada pelo Plasmodium chabaudi. O efeito sobre o nosso velho conhecido Anopheles gambiae, que transmite ao homem o Plasmodium falciparum, é estudado no mesmo número da Science num artigo (ref2) de Ernst-Jan Scholte e colegas. Numa tradução livre do resumo:O controlo biológico de mosquitos da malária em África tem sido usado raramente em programas de controlo de vectores. Desenvolvimentos recentes neste ramo mostram que certos fungos são virulentos para mosquitos Anopheles adultos. A pulverização de forma práctica de um fungo entomopatogénico que infectou e matou Anopheles gambiae adultos, o principal vector da malária em África, foi conseguida em casas rurais africanas. Um modelo de inoculação entomológico sugere que a implementação deste método de controlo do vector, mesmo com a cobertura moderada observada durante um estudo de campo na Tânzania, iria reduzir a intensidade de transmissão da malária de forma significativa.
O estudo tem duas partes. Na primeira os autores verificaram que em cinco casas em que colocaram fungos durante 5 semanas, cerca de 23% dos mosquitos fêmeas foram infectadas pelo fungo. Com base num modelo matemático verificaram que isso significaria que num ano o número de picadas potencialmente infecciosas por aldeão baixaria de 262 em média para 64. Os números poderiam baixar para qualquer coisa em torno de uma picada por ano, valor necessário para realmente combater a malária, se se procedesse à pulverização de paredes inteiras em todas as casas da aldeia. E nestes cálculos não entraram em conta com os efeitos sobre os mosquitos infestados pelo plasmódio discutidos no artigo de Blanford e colegas.
Os fungos são particularmente interessantes pois não se conhecem casos em que os insectos tenham desenvolvido resistência a este tipo de agentes. Por outro lado a infecção é muito simples, o menor contacto de um esporo de um fungo com a pata de um mosquito é suficiente para despoletar a infecção, não sendo necessário ingeri-los. Estes dois estudos abrem possibilidades interessantes, quer pela utilização do fungo como agente principal, quer combinado com pesticidas químicos, pois é pouco provável que os mosquitos consigam desenvolver resistência a ambos. Qualquer passo no combate a uma doença que mata uma criança africana a cada 30 segundos é importante, e voltarei a este tema após o feriado do dia 1 de Maio.
Referências
(ref1) Simon Blanford, Brian H. K. Chan, Nina Jenkins, Derek Sim, Ruth J. Turner, Andrew F. Read, Matt B. Thomas (2005). Fungal Pathogen Reduces Potential for Malaria Transmission. Science, Vol. 308. no. 5728, pp. 1638 - 1641. Laço DOI
(ref2) Ernst-Jan Scholte, Kija Ng'habi, Japheth Kihonda, Willem Takken, Krijn Paaijmans, Salim Abdulla, Gerry F. Killeen, Bart G. J. Knols (2005). An Entomopathogenic Fungus for Control of Adult African Malaria Mosquitoes. Science, Vol. 308. no. 5728, pp. 1641 - 1642. Laço DOI
sábado, abril 29, 2006
O mosquito e o fungo que o mata
sexta-feira, abril 28, 2006
Maus como as mães


 Quão diferentes poderiam ser as coisas se as mulheres tivessem tido um papel mais dominante na história humana? É possível que não fossem tão diferentes como isso. Com efeito, existe na natureza um exemplo de uma sociedade de predadores dominada pelas fêmeas da espécie. Trata-se da hiena malhada, de nome científico Crocuta crocuta. Um conjunto de características únicas fazem destes animais um objecto de estudo fascinante. As hienas malhadas vivem em grupos sociais que podem incluir 40 a 60 indivíduos, com uma estrutura hierárquica bastante rígida, e uma organização de tipo quase militar. A sociedade das hienas faz lembrar em muitos aspectos grupos de primatas como os dos babuínos, mas com uma inversão no estatuto de machos e fêmeas. [... ler mais]
Quão diferentes poderiam ser as coisas se as mulheres tivessem tido um papel mais dominante na história humana? É possível que não fossem tão diferentes como isso. Com efeito, existe na natureza um exemplo de uma sociedade de predadores dominada pelas fêmeas da espécie. Trata-se da hiena malhada, de nome científico Crocuta crocuta. Um conjunto de características únicas fazem destes animais um objecto de estudo fascinante. As hienas malhadas vivem em grupos sociais que podem incluir 40 a 60 indivíduos, com uma estrutura hierárquica bastante rígida, e uma organização de tipo quase militar. A sociedade das hienas faz lembrar em muitos aspectos grupos de primatas como os dos babuínos, mas com uma inversão no estatuto de machos e fêmeas. [... ler mais]
Um artigo sobre estas criaturas acaba de sair na Nature (ref1) da autoria de Dloniak e colegas, e eu não resisto a discuti-lo aqui. A imagem ao lado mostra exactamente a primeira autora do estudo a pesar uma das hienas. Numa tradução livre de um excerto da introdução do artigo:A sociedade da hiena malhada é estruturada numa hierarquia de dominância rígida, e a posição social do indivíduo determina a sua prioridade no acesso aos recursos, bem como o seu sucesso reprodutivo. Num grupo social, as hienas fêmeas variam por isso consideravelmente no que respeita a condição, dependendo do estatuto social que herdam das suas mães. As fêmeas das hienas malhadas são geralmente maiores que os machos, e exibem genitais fortemente masculinizados através dos quais o parto é dificílimo. As fêmeas são também mais agressivas que os machos e socialmente dominantes em relação a eles.
 A submissão que os machos parecem aceitar de forma tão surpreendente tem custos mas também benefícios:
A submissão que os machos parecem aceitar de forma tão surpreendente tem custos mas também benefícios: Em contraste com a maioria dos machos dos primatas, as hienas macho observadas fora do seu território natal comportam-se de forma submissa face a cada membro da mesma espécie que encontrem, qualquer que seja o seu sexo, idade, ou tamanho corporal relativo. Como resultado, os machos caem inevitavelmente na hierarquia quando se dispersam e sofrem um declínio substancial na prioridade de acesso ao alimento aquando do abate de ungulados. Contudo, apesar desse custos energéticos da emigração, os machos raramente acasalam a menos que dispersem. Logo, de um ponto de vista adaptativo, os machos parecem "trocar" comida por sexo quando dispersam dos seus clãs natais.
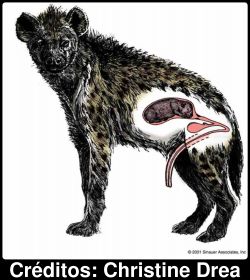 Os genitais das hienas malhadas fêmeas são de facto tão semelhantes aos dos machos que Aristóteles, por exemplo, pensava que as hienas eram hermafroditas. A ilustração ao lado mostra uma fêmea grávida com alguns detalhes do aparelho reprodutor feminino visíveis. O tubo, que atinge cerca de 15 cm de comprimento fora do corpo, não se trata de um pénis, embora se assemelhe: esta é na verdade a forma que tomam os orgão genitais femininos. Não custa a imaginar as dificuldades que algo tão peculiar representa para a cópula, e sobretudo para o parto. Por incrível que pareça, para nascerem as pequenas hienas têm que percorrer toda a estrutura. As dificuldades em fazer passar duas crias através de um tubo que, mesmo após alargar para o dobro do diâmetro, fica apenas com cerca de cinco centímetros da largura, são enormes, e muitas fêmeas morrem ao tentar dar à luz a primeira ninhada. É óbvio que se esta masculinização das hienas acarreta custos elevados, também deve acarretar benefícios.
Os genitais das hienas malhadas fêmeas são de facto tão semelhantes aos dos machos que Aristóteles, por exemplo, pensava que as hienas eram hermafroditas. A ilustração ao lado mostra uma fêmea grávida com alguns detalhes do aparelho reprodutor feminino visíveis. O tubo, que atinge cerca de 15 cm de comprimento fora do corpo, não se trata de um pénis, embora se assemelhe: esta é na verdade a forma que tomam os orgão genitais femininos. Não custa a imaginar as dificuldades que algo tão peculiar representa para a cópula, e sobretudo para o parto. Por incrível que pareça, para nascerem as pequenas hienas têm que percorrer toda a estrutura. As dificuldades em fazer passar duas crias através de um tubo que, mesmo após alargar para o dobro do diâmetro, fica apenas com cerca de cinco centímetros da largura, são enormes, e muitas fêmeas morrem ao tentar dar à luz a primeira ninhada. É óbvio que se esta masculinização das hienas acarreta custos elevados, também deve acarretar benefícios.
Para tentarem perceber melhor os mecanismos que levam a esta masculinização das fêmeas das hienas os autores resolveram estudar os níveis hormonais das diferentes fêmeas, em particular durante os períodos de gravidez.Excepto durante a gravidez, as hienas fêmeas têm níveis de testosterona no sangue inferiores aos dos machos adultos, indicando que os efeitos dos andrógenos não explicam a diferença de agressividade em função do sexo documentada na espécie. Contudo, os ovários de hienas fêmeas grávidas produzem grandes quantidades de hormonas esteróides andrógenas durante a segunda metade da gestação, e estas atravessam a placenta atingindo o feto em desenvolvimento. Logo, as crias de hiena de ambos os sexos são expostas a altas concentrações de andrógenos durante a vida fetal.
A forma como os autores se propuseram estudar isto pode parecer pouco digna mas foi eficaz.Testamos aqui a hipótese de que a exposição pré-natal das crias a andrógenos afecta o fenótipo da descendência nas hienas. Investigamos em primeiro lugar os padrões de concentração de andrógenos fecais ao longo da gestação em fêmeas grávidas.
OS autores recolheram 53 amostras fecais de um total de 43 gravidezes de 27 hienas fêmeas adultas, que foram congeladas, guardadas e posteriormente analisadas. Não deixa de ser estranho pensar que após tantos anos de faculdade, mestrados e doutoramentos, o emprego destes cientistas consista em grande parte em seguir hienas e recolher os seus excrementos. Os resultados da análise são descritos no que se segue.Mostramos que as concentrações de andrógenos em hienas malhadas fêmeas (Crocuta crocuta) são mais elevadas durante a fase tardia da gestação em fêmeas dominantes que em fêmeas subordinadas. Para além disso, quer as crias machos quer as crias fêmeas nascidas de mães com concentrações mais elevadas de andrógenos na fase tardia da gravidez exibem taxas de agressividade mais elevadas e comportamento de "montar" que crias nascidas de mães com concentrações de andrógenos mais baixas.
A sugestão dos autores é que a vantagem tem a ver com os hábitos alimentares das hienas.Os nossos resultados oferecem o primeiro laço definitivo entre andrógenos pré-natais e agressividade em fêmeas de hienas malhadas, e apoiam a hipótese que a selecção para agressividade debaixo de uma intensa competição para se alimentar foi a pressão selectiva inicial que levou ao aumentar da agressividade da agressividade e de outras características masculinizadas das hienas fêmeas. A agressividade nas hienas malhadas fêmeas deve ser adaptativa porque a competição pelo alimento nesta espécie é muito intensa, e as fêmeas mais capazes de desalojar os congéneres do acesso ao alimento nos locais de abate de ungulados obtêm o sucesso reprodutivo mais elevado.

Há um outro aspecto mais curioso que os autores focam.A dominação social e a masculinização das fêmeas tornam o sexo extremamente desafiador para as hienas macho, pelo que machos que capacidade de acasalamento mais elevada deverão ser favorecidos pela seleção natural. Orientar correctamente a fêmea durante o acto de montar é crítico para o macho conseguir a intromissão durante o acto sexual, e a quantidade de práctica que o macho consegue cedo na vida pode afectar profundamente o seu sucesso reprodutivo.
Na verdade dada a anatomia dos orgãos genitais femininos a tarefa dos machos hiena afigura-se-me particularmente difícil, e percebo que precisem de treinar bastante. Infelizmente não consegui nenhuma imagem que ilustrasse o acto. Se algum dos leitores me conseguir arranjar pornografia de Crocuta crocuta desde já agradeço. Como despedida eis mais uma imagem destes seres, que embora tão agressivos, são mães extremosas.
Ficha técnica
As imagens utlizadas nesta contribuição, bem como alguma da informação, foram retiradas da página de Kay Holemkamp, co-autora do artigo indicado como ref1 abaixo.
Referências
(ref1) S. M. Dloniak, J. A. French and K. E. Holekamp (2006). Rank-related maternal effects of androgens on behaviour in wild spotted hyaenas. Nature 440, 1190-1193. Laço DOI.
quinta-feira, abril 27, 2006
Chimpanzés cidadãos de Espanha

 Uma certa sobranceria é normal entre vizinhos e dizer mal das pessoas do outro lado da fronteira é um daqueles pequenos prazeres que, se bem que mesquinho, a maioria das pessoas aprecia secretamente, mesmo que se mostrem escandalizados e finjam que não. E confesso que descobri hoje que também sou culpado disso. Quando li no John Hawks Anthropology Weblog que os espanhóis se preparavam para votar uma lei para reconhecer aos grandes símios direitos iguais aos dos seres humanos pensei logo para comigo: só mesmo em Espanha. Mas depois perguntei-me se a notícia não seria uma piada, talvez o dia das mentiras seja mais tarde no país vizinho. [... ler mais]
Uma certa sobranceria é normal entre vizinhos e dizer mal das pessoas do outro lado da fronteira é um daqueles pequenos prazeres que, se bem que mesquinho, a maioria das pessoas aprecia secretamente, mesmo que se mostrem escandalizados e finjam que não. E confesso que descobri hoje que também sou culpado disso. Quando li no John Hawks Anthropology Weblog que os espanhóis se preparavam para votar uma lei para reconhecer aos grandes símios direitos iguais aos dos seres humanos pensei logo para comigo: só mesmo em Espanha. Mas depois perguntei-me se a notícia não seria uma piada, talvez o dia das mentiras seja mais tarde no país vizinho. [... ler mais]
Após uma busca na Internet encontrei alguns laços e blogs que discutiam a notícia. Traduzida a coisa dá:Com a justificação de o homem partilhar 98.4% dos genes com os chimpanzés, 97.7% com os gorilas e 96.4% com os orangutangos, o grupo socialista apresentará um projecto que pede "direitos humanos" para os símios. Uma organização internacional com o mesmo nome do projecto procura uma declaração da ONU sobre os direitos dos símios e defende direitos iguais aos dos "menores de idade e aos incapacitados mentais da nossa espécie", segundo os responsáveis do projecto.
O ênfase é meu, e levou-me a desconfiar ainda mais da notícia, se bem que a imprensa manobre muitas vezes as notícias colocando as palavras fora do contexto, para conseguir mais sensacionalismo. A fonte pode ser encontrada aqui, ou aqui. Mas tudo parece ser especulação mediática desenfreada. Há de facto um projecto de lei sobre maus-tratos e utilização de grandes símios em espectáculos de circo e experiências. Depois de procurar um bocado encontrei um artigo que me pareceu corresponder melhor à verdade dos factos:A ministra do meio ambiente, Cristina Narbona, assegurou hoje que a proposta de lei apresentada pelo Proyecto Gran Simio como o apoio do PSOE não pretende em caso algum reconhecer "direitos humanos" a estes animais desmentindo o sentido com que algumas informações o interpretaram. "Isso não está correcto, houve uma interpretação equivocada", salientou.
E mais adiante"Em caso algum se criam ou se defendem direitos humanos para os símios; em nenhum momento se fala de outorgar direitos humanos aos grandes símios", reiterou.
Mais uma vez o ênfase é meu. As citações foram encontradas aqui na cadenaser. Embora este artigo não seja claro, pois depois volta a bater na tecla dos direitos humanos, e fala mesmo de uma reacção da igreja, esta história parece ser um caso de orgãos de comunicação social a tentarem arranjar controvérsia para vender papel. Nada a que não estejamos habituados deste lado da fronteira.
terça-feira, abril 25, 2006
Escondida do registo fóssil

 Esta criatura que parece um camarão algo estranho, o Neoglyphea neocaledonica, é uma nova espécie, recém-descoberta. A descoberta de uma nova espécie, embora seguramente sempre emocionante para o descobridor, não é tão rara como isso, basta ver as rãs que têm sido descobertas na selva do Laos nos últimos tempos. O Neoglyphea é algo diferente, trata-se de um "fóssil vivo", isto é um membro de um grupo de animais cujos últimos vestígios fósseis datam de há muitos milhões de anos. O Neoglyphea representa para os estudiosos dos crustáceos o que a descoberta do celacanto, Latimeria chalumnae, representou para os especialistas dos vertebrados. O antepassado de todos os decápodes actuais (caranguejos, lagostas e camarões) pertenceria ao grupo dos glifeídeos. [... ler mais]
Esta criatura que parece um camarão algo estranho, o Neoglyphea neocaledonica, é uma nova espécie, recém-descoberta. A descoberta de uma nova espécie, embora seguramente sempre emocionante para o descobridor, não é tão rara como isso, basta ver as rãs que têm sido descobertas na selva do Laos nos últimos tempos. O Neoglyphea é algo diferente, trata-se de um "fóssil vivo", isto é um membro de um grupo de animais cujos últimos vestígios fósseis datam de há muitos milhões de anos. O Neoglyphea representa para os estudiosos dos crustáceos o que a descoberta do celacanto, Latimeria chalumnae, representou para os especialistas dos vertebrados. O antepassado de todos os decápodes actuais (caranguejos, lagostas e camarões) pertenceria ao grupo dos glifeídeos. [... ler mais]
Esta é a segunda espécie a ser descoberta neste género. Foi capturada em Outubro de 2005 numa expedição liderada por Bertrand Richer de Forges e é descrita na revista Zoosytema (ref1). Numa tradução livre do resumo original em francês:Uma nova espécie de glifeídeo do género até ao presente monotípico Neoglyphea, N. neocaledonica foi capturada nas águas profundas do mar do Coral. Após uma descrição do único exemplar feminino, uma comparação morfológica é feita com a outra espécie do género, N. inopinata.
O artigo é sobretudo a descrição minuciosa da morfologia da criatura e da aspectos filogenéticos. Aqueles que consigam ler francês fluentemente poderão consultar o pdf do artigo, que se encontra disponível para consulta na internet. Não resisto aos detalhes sobre a descoberta da primeira espécie:Quando em 1975 dois investigadores, J. Forest et M. de Saint Laurent, mencionaram que um crustáceo recolhido pelo navio de pesquisa americana Albatroz nas Filipinas, em 1908, a 186 metros de profundidade e conservado depois, não identificado, no Museu de História Natural de Washington, pertencia ao grupo dos glifeídeos conhecido do Mesozóico e do qual se supunham todos os representantes desaparecidos desde o Cretácico Superior, a notícia provocou um grande alarido. Esse "fóssil vivo" foi então descrito com o nome Neoglyphea inopinata.
O ênfase a negrito é meu. Os antepassados do animal não só conseguiram esconder-se do registo fóssil durante cerca de 50 milhões de anos, como ainda este exemplar permaneceu à vista de toda a gente sem ser identificado durante 60 anos após ser capturado. Realmente a criatura não tinha vontade de ser "descoberta" pelos seres humanos. Os 50 milhões de anos vêm do comunicado de imprensa do CenSeam onde se refere que se pensava que os glifeídeos teriam desaparecido no Eoceno. Quanto ao N. neocaledonica:Apesar da sua recolha em águas profundas, o animal pôde ser observado vivo. É muito activo e agressivo e utiliza a sua pseudo-pinça para atacar, apoiando-se sobre o primeiro conjunto de patas ambulatórias, com o cefalotórax formando uma curva com o abdómen.
Como vemos o N. neocaledonica também não parece muito feliz por ter sido descoberto. A atenção para este artigo chegou-me através do blog Deep-Sea News, o local por excelência para descobrir notícias recentes sobre as profundezas marinhas.
Referências
(ref1)
sábado, abril 22, 2006
Grilos suicidas e os parasitas que não querem ser comidos

 O grilo da figura está tranquilamente a cuidar da sua vida quando, de repente, num aparente acesso de loucura suicida, corre para a borda de uma piscina e se atira lá para dentro. Mas as coisas não são bem assim. Trata-se de mais um tenebroso exemplo daquilo que os parasitas levam os seus hospedeiros a fazer. Exemplos destes já foram aqui discutidos, em particular o caso da Ampulex compressa e do Toxoplasma gondii. Hoje o vilão é o verme górdio, de seu nome científico Paragordius tricuspidatus. [... ler mais]
O grilo da figura está tranquilamente a cuidar da sua vida quando, de repente, num aparente acesso de loucura suicida, corre para a borda de uma piscina e se atira lá para dentro. Mas as coisas não são bem assim. Trata-se de mais um tenebroso exemplo daquilo que os parasitas levam os seus hospedeiros a fazer. Exemplos destes já foram aqui discutidos, em particular o caso da Ampulex compressa e do Toxoplasma gondii. Hoje o vilão é o verme górdio, de seu nome científico Paragordius tricuspidatus. [... ler mais] Durante a fase inicial da sua vida o verme górdio parasita os grilos, mas durante a fase adulta vive livremente na água, onde se reproduz. Daí o parasita manipular o comportamento dos grilos e levá-los a adoptar o comportamento suicida descrito acima. Uma vez dentro de água não deixa de ser impressionante ver o tamanho do verme que sai de dentro do grilo. Ora devido a esse tamanho o verme demora um certo tempo a sair: a tarefa pode levar qualquer coisa como 10 minutos. Durante esse tempo os movimentos do grilo e do verme a tentar sair do seu hospedeiro podem atrair predadores do grilo, tais como rãs ou peixes. O verme górdio não parasita nenhum desses animais e poderia pensar-se que se o grilo for comido então é também o fim do verme. Pelos vistos nem sempre é assim. Num artigo da Nature (ref1), Fleur Ponton e colegas descrevem o que sucede quando grilos infestados com o Paragordius tricuspidatus são ingeridos por predadores. Numa tradução livre do resumo:
Durante a fase inicial da sua vida o verme górdio parasita os grilos, mas durante a fase adulta vive livremente na água, onde se reproduz. Daí o parasita manipular o comportamento dos grilos e levá-los a adoptar o comportamento suicida descrito acima. Uma vez dentro de água não deixa de ser impressionante ver o tamanho do verme que sai de dentro do grilo. Ora devido a esse tamanho o verme demora um certo tempo a sair: a tarefa pode levar qualquer coisa como 10 minutos. Durante esse tempo os movimentos do grilo e do verme a tentar sair do seu hospedeiro podem atrair predadores do grilo, tais como rãs ou peixes. O verme górdio não parasita nenhum desses animais e poderia pensar-se que se o grilo for comido então é também o fim do verme. Pelos vistos nem sempre é assim. Num artigo da Nature (ref1), Fleur Ponton e colegas descrevem o que sucede quando grilos infestados com o Paragordius tricuspidatus são ingeridos por predadores. Numa tradução livre do resumo:Como prisioneiros no seu habitat vivo os parasitas são vulneráveis à destruição do seu hospedeiro por predadores. Contudo, mostramos aqui que o verme górdio parasita, Paragordius tricuspidatus é capaz não apenas de escapar de dentro do insecto seu hospedeiro após a ingestão por um peixe ou rã, mas também do aparelho digestivo do predador. Esta estratégia notável permite ao verme prosseguir com o seu ciclo de vida.
Isto pode ser observado abaixo em que se mostra uma rã que acabou de engolir um infeliz grilo. Na imagem da direita pode já ver-se um verme a tentar escapar para a liberdade. Apesar dos esforços da rã para não deixar fugir a criatura o verme lá consegue a sua liberdade, e vive para se ir juntar a outros vermes na água.
Apesar dos esforços da rã para não deixar fugir a criatura o verme lá consegue a sua liberdade, e vive para se ir juntar a outros vermes na água.
Pequenos filmes com a corrida suicida dos grilos, vermes a escaparem da boca das rãs e das guelras de peixes podem ser encontrados nas páginas de material suplementar do artigo na Nature. Os autores do estudo observaram 477 pobres grilos que foram comidos por uma variedade de predadores após se atirarem para a água. Em nenhum desses casos os predadores cuspiram o grilo, mas uma fracção significativa dos vermes conseguiu escapar: entre 18% a 26% dos ingeridos por várias espécies de peixes, e 35% dos ingeridos pelas rãs. Os autores verificaram que se o verme não aparecesse na boca, narinas ou guelras nos cinco minutos a seguir a ser ingerido não conseguia escapar, morrendo provavelmente no estômago do predador do grilo. O tempo de fuga era em média de 8 minutos e meio mas podia ser substancialmente maior pois os predadores tentavam engoli-los novamente. O mais longo observado foram 28 minutos,
Para lá do aspecto repugnante e nojento da história que eu pessoalmente aprecio, o estudo mostra como um parasita pode eventualmente desenvolver um ciclo de vida complexo. Tudo pode começar por tentar sobreviver à ingestão acidental do seu hospedeiro, levando progressivamente o parasita a incluir o predador no seu ciclo de vida. Eu realmente adoro histórias de parasitas, especialmente estas, com um final feliz. Assim, para terminar, nada melhor que mais umas fotos do parasita em toda a sua glória, a escapar rumo à liberdade.
Referências
(ref1) Fleur Ponton, Camille Lebarbenchon, Thierry Lefèvre, David G. Biron, David Duneau, David P. Hughes and Frédéric Thomas (2006). Parasitology: Parasite survives predation on its host. Nature 440, 756. Laço DOI.
quinta-feira, abril 20, 2006
As pernas da serpente

 Esta imagem mostra parte do fóssil da Najash rionegrina, uma cobra que viveu há cerca de 90 milhões de anos na região que hoje em dia é a Patagónia na Argentina. Quando se pensa em cobra, a imagem que vem à mente é um animal rastejante, sem membros. Mas não foi sempre assim, e não é por acaso que o nome genérico desta criatura vem do hebraico, e designa a serpente com pernas que aparece no início da bíblia. Aqueles com algum conhecimento da anatomia dos vertebrados não terão dificuldade em identificar os ossos da pernas entre estes vestígios. [... ler mais]
Esta imagem mostra parte do fóssil da Najash rionegrina, uma cobra que viveu há cerca de 90 milhões de anos na região que hoje em dia é a Patagónia na Argentina. Quando se pensa em cobra, a imagem que vem à mente é um animal rastejante, sem membros. Mas não foi sempre assim, e não é por acaso que o nome genérico desta criatura vem do hebraico, e designa a serpente com pernas que aparece no início da bíblia. Aqueles com algum conhecimento da anatomia dos vertebrados não terão dificuldade em identificar os ossos da pernas entre estes vestígios. [... ler mais]
Conheciam-se já três espécies fósseis de cobras com membros no exterior do corpo, e todos eles eram vestígios de cobras marinhas. Devido a esse facto admitia-se como provável que a anatomia das cobras seria uma adaptação a um estilo de vida aquático. Ora a descoberta da Najash mudou tudo isso. Os restos fósseis da Najash são descritos num artigo na Nature da autoria de Sebastián Apesteguía e Hussam Zaher, este último do Museu de Zoologia da Universidade de São Paulo no Brasil. Numa tradução livre do resumo:Era habitual pensar que as serpentes perderam os seus membros de forma progressiva devido a uma diminuição gradual da sua utilização. Contudo descobertas de desenvolvimento e paleontológicas sugerem um cenário mais complexo para a redução dos membros, ainda que documentado de forma pobre no registo fóssil. Relatamos aqui uma cobra fóssil com um sacro suportando uma cintura pélvica e pernas robustas e funcionais fora da caixa torácica. O novo fóssil, do Cretácico Superior da Patagónia, preenche um hiato importante na progressão evolutiva no sentido da perda de membros pois outras cobras fósseis conhecidas com membros posteriores desenvolvidos, as cobras marinhas Haasiophis, Pachyrhachis e Eupodophis não possuem a região do sacro.
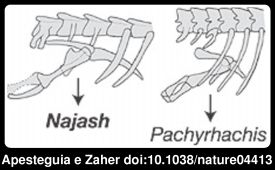 A região que destaquei a negrito é um pormenor muito importante. Tal como se mostra na imagem ao lado, as pernas da Najash ainda articulam com a coluna vertebral, enquanto nos fósseis das outras cobras com pernas isso não se passa. Isso significa que a Najash representa um estágio evolutivo "mais primitivo" que aquele encontrado nas cobras marinhas. O fóssil é razoavelmente completo, com material craniano que, juntamente com outras características do esqueleto, apoiam essa hipótese.
A região que destaquei a negrito é um pormenor muito importante. Tal como se mostra na imagem ao lado, as pernas da Najash ainda articulam com a coluna vertebral, enquanto nos fósseis das outras cobras com pernas isso não se passa. Isso significa que a Najash representa um estágio evolutivo "mais primitivo" que aquele encontrado nas cobras marinhas. O fóssil é razoavelmente completo, com material craniano que, juntamente com outras características do esqueleto, apoiam essa hipótese.Uma análise filogenética mostra que o novo fóssil é a cobra mais primitiva (basal) conhecida e que todas as outras cobras com membros conhecidas estão mais próximas das serpentes macróstomas, um grupo que inclui as boas, pitões e colubróides.
Ora a partir do esqueleto os autores conseguem inferir o modo de vida da Najash, e sugerem um cenário alternativo para a origem das cobras.O novo fóssil mantêm certas características associadas com um modo de vida subterrânea, também presentes em linhagens mais primitivas de serpentes existentes hoje em dia, apoiando a hipótese de uma origem terrestre em vez de marinha para as cobras.
Deve notar-se que as cobras não são os únicos tetrápodes ápodes (sem membros). Existe, por exemplo, um grupo de anfíbios, as cecílias, de que falámos aqui recentemente, e um outro grupo de répteis, os anfisbenídeos, que também perderam os seus membros. Nesses dois casos tratou-se de uma adaptação a um modo de vida fossorial, isto é subterrâneo. Esta nova descoberta, embora não resolva de vez a questão, aponta no mesmo sentido. São no entanto precisos mais fósseis, pois o registo fóssil da cobras, que evoluiram a partir de um animal aparentado aos lagartos monitores, há cerca de 145 milhões de anos, é muito pobre. Não deixa de ser curioso que, depois de se falar acerca da evolução das pernas a partir das barbatanas dos peixes, aquando do tiktaalik, se fale agora numa descoberta sobre a evolução em sentido inverso.
Uma visão artística do aspecto da cobra pode ser encontrada aqui nas páginas da LiveScience. As pernas apesar de robustas e funcionais são minúsculas, e a utilização que a Najash dava aos seus membros não é conhecida.
Referências
(ref1) Sebastián Apesteguía, and Hussam Zaher (2006). A Cretaceous terrestrial snake with robust hindlimbs and a sacrum. Nature 440, 1037-1040. Laço DOI
quarta-feira, abril 19, 2006
Como esticar os dedos para fazer uma asa
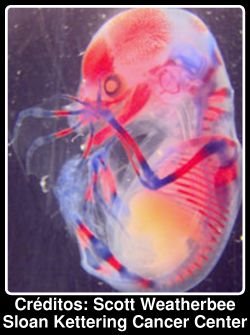
 Esta criatura com ar sinistro é um embrião de um morcego com 80 dias. Com esta idade já é visível a característica mais marcante destes animais: os extraordinariamente longos dedos das mãos. Os morcegos são uma história de sucesso entre os mamíferos: uma em cada cinco espécies de mamíferos conhecidos é uma espécie de morcego. Estas criaturas evoluiram muito provavelmente de animais semelhantes a musaranhos, mas a forma como ocorreu essa evolução não é conhecida, pois não foram encontradas quaisquer formas intermédias. O registo fóssil dos morcegos mostra que as primeiras formas, há cerca de 50 milhões de anos, tinham asas essencialmente iguais às formas modernas. Isto sugere uma evolução muito rápida, quase súbita, das asas. [... ler mais]
Esta criatura com ar sinistro é um embrião de um morcego com 80 dias. Com esta idade já é visível a característica mais marcante destes animais: os extraordinariamente longos dedos das mãos. Os morcegos são uma história de sucesso entre os mamíferos: uma em cada cinco espécies de mamíferos conhecidos é uma espécie de morcego. Estas criaturas evoluiram muito provavelmente de animais semelhantes a musaranhos, mas a forma como ocorreu essa evolução não é conhecida, pois não foram encontradas quaisquer formas intermédias. O registo fóssil dos morcegos mostra que as primeiras formas, há cerca de 50 milhões de anos, tinham asas essencialmente iguais às formas modernas. Isto sugere uma evolução muito rápida, quase súbita, das asas. [... ler mais]
A ausência de fósseis não desencorajou os cientistas, que num artigo recente nos Proceedings of the National Academy of Sciences (ref1), da autoria de Karen E. Sears e colegas, recorrem ao estudo de embriões de ratos e de morcegos para descobrirem as diferenças na expressão dos genes que controlam o desenvolvimento dos dedos das mãos de ratos e morcegos, e que são também responsáveis pelas diferenças entre os dedos dos pés e das mãos dos morcegos. Paradoxalmente tudo parece repousar na expressão de uma única proteína. Numa tradução livre do resumo:Os mais antigos fósseis de morcegos assemelham-se às formas modernas ao possuirem dedos muito alongados para suportar a membrana da asa, que é a marca anatómica do vôo propulsionado. Para confirmar quantitativamente estas semelhanças, executámos uma análise morfométrica de ossos da asas de morcegos modernos e de morcegos fósseis. Verificámos que os comprimentos dos terceiro, quarto e quinto dedos (os elementos mais importantes no suporte da asa) permaneceram constantes relativamente ao tamanho do corpo nos últimos 50 milhões de anos. Esta ausência de formas transicionais no registo fóssil levou-nos a procurar noutros locais para compreender a evolução da asa dos morcegos.
Os "outro local" de que os autores falam é o desenvolvimento embrionário, Como os morcegos descendem de animais que teriam mãos semelhantes às dos ratos, os autores resolveram comparar o desenvolvimento relativo nos embriões de ratos (20 dias) e morcegos (120 dias). Como referem no que se segue, o mecanismo de diferenciação é simples:Ao investigar o desenvolvimento embrionário, verificámos que os dedos nos morcegos (Carollia perspicillata) são inicialmente semelhantes em comprimento aos dos ratos (Mus musculus) mas que, subsequentemente, os dedos do morcegos se alongam grandemente. O momento do desenvolvimento da mudança no comprimento dos dedos aponta para uma mudança na cartilagem longitudinal, um processo que depende da proliferação e diferenciação dos condrócitos. Verificámos que os dedos dos membros anteriores dos morcegos exibem taxas relativamente elevadas de proliferação e diferenciação de condrócitos. Mostramos que a proteína morfogenética do osso (Bmp2) pode estimular a proliferação, diferenciação e aumento no comprimento dos dedos nos membros anteriores dos embriões dos morcegos.
Esta mudança nos padrões de crescimento entre as mãos dos ratos e morcegos ocorre a meio do período de gestação. Antes disso os membros anteriores de ratos e morcegos são virtualmente idênticos, como se pode ver na imagem abaixo (o embrião do rato tem cerca de 12 dias e meio).
Durante o período em que os dedos das mãos dos morcegos "esticam", os dedos dos pés dos morcegos e ratos mostram desenvolvimento idêntico. Os autores sugerem que a diferença observada nas mãos tem a ver com um acréscimo de cerca de 35% na expressão de um factor de crescimento do tecido ósseo (a proteína Bmp2). Além disso, mostramos que a expressão e a sinalização de Bmp2 são aumentadas no desenvolvimento embrionário dos dedos nos membros anteriores dos morcegos relativamente aos dedos nos membros posteriores dos ratos ou morcegos. Considerados em conjunto, os nossos resultados sugerem que uma regulação a nível superior do ciclo do Bmp2 é um dos factores mais importantes no desenvolvimento do alongamento dos dedos dos membros anteriores dos morcegos, e que é também potencialmente um mecanismo chave no seu elongamento evolutivo.
Trata-se de uma pesquisa que já tinha sido apresentada em congressos em 2004, e é agradável ver que finalmente foi publicada. Como vemos a parte da evolução das asas parece relativamente simples e isso pode explicar a rapidez com que os morcegos aparecem no registo fóssil. O pequeno acréscimo de 35% no regulador de crescimento é provavelmente o que tornou possível o vôo com os dedos das mãos 50 milhões de anos atrás.
Referências
(ref1) Karen E. Sears, Richard R. Behringer, John J. Rasweiler IV, and Lee A. Niswander (2006). Development of bat flight: Morphologic and molecular evolution of bat wing digits. Proc. Natl. Acad. Sci. USA. Laço DOI.
quinta-feira, abril 13, 2006
Tomai meus filhos, comei da minha carne

 Não se trata de nenhuma incursão por terrenos religiosos, nem tem a ver com o feriado religioso que atravessamos. A criatura de hoje é uma mãe que vai ao extremo de alimentar os filhos com a própria carne. A coisa com aspecto de minhoca que se vê ao lado, a Boulengerula taitanus, é na verdade um vertebrado, de um grupo de anfíbios designados por cecílias. As cecílias têm hábitos fossoriais e são predadores eficazes que vivem nas zonas húmidas de África e América do Sul. No Brasil são chamadas de cobras-cegas, e assemelham-se às "cobras-cegas" que se encontram em Portugal, embora não sejam a mesma coisa. As cobras cegas portuguesas são répteis, mais exactamente anfisbenídeos, e a parecença é resultado da convergência em função de um modo de vida semelhante. A espécie Boulengerula taitanus intrigou os cientistas devido ao facto de as fêmeas com crias apresentarem um tom de pele diferente das fêmeas sem crias. [... ler mais]
Não se trata de nenhuma incursão por terrenos religiosos, nem tem a ver com o feriado religioso que atravessamos. A criatura de hoje é uma mãe que vai ao extremo de alimentar os filhos com a própria carne. A coisa com aspecto de minhoca que se vê ao lado, a Boulengerula taitanus, é na verdade um vertebrado, de um grupo de anfíbios designados por cecílias. As cecílias têm hábitos fossoriais e são predadores eficazes que vivem nas zonas húmidas de África e América do Sul. No Brasil são chamadas de cobras-cegas, e assemelham-se às "cobras-cegas" que se encontram em Portugal, embora não sejam a mesma coisa. As cobras cegas portuguesas são répteis, mais exactamente anfisbenídeos, e a parecença é resultado da convergência em função de um modo de vida semelhante. A espécie Boulengerula taitanus intrigou os cientistas devido ao facto de as fêmeas com crias apresentarem um tom de pele diferente das fêmeas sem crias. [... ler mais] Para além disso, as crias desta espécie apresentam uma boca cheia de dentes, que se mostra na imagem abaixo.
Para além disso, as crias desta espécie apresentam uma boca cheia de dentes, que se mostra na imagem abaixo.
 O pequeno traço que define a escala no canto inferior direito corresponde a 30 micrómetros. Estes dentes são bastante diferentes dos dentes dos adultos, parecendo-se com pequenos ganchos, como se pode ver na ampliação à direita. Outras espécies de cecílias têm dentes especializados enquanto jovens, mas essas espécies são vivíparas. Nessas espécies os dentes são utilizados pelas crias para rasparem o oviduto da mãe, que possui uma camada especial de células das quais as crias se alimentam (ref1). Ora a Boulengerula taitanus é uma espécie ovípara, e os cientistas pensaram que, já que as crias não comiam a carne da mãe a partir do interior, talvez o fizessem do exterior. Para isso recolheram alguns exemplares que estudaram no laboratório. O resultado desse estudo de Alexander Kupfer e colegas é descrito num artigo na Nature (ref2). Numa tradução livre do resumo:
O pequeno traço que define a escala no canto inferior direito corresponde a 30 micrómetros. Estes dentes são bastante diferentes dos dentes dos adultos, parecendo-se com pequenos ganchos, como se pode ver na ampliação à direita. Outras espécies de cecílias têm dentes especializados enquanto jovens, mas essas espécies são vivíparas. Nessas espécies os dentes são utilizados pelas crias para rasparem o oviduto da mãe, que possui uma camada especial de células das quais as crias se alimentam (ref1). Ora a Boulengerula taitanus é uma espécie ovípara, e os cientistas pensaram que, já que as crias não comiam a carne da mãe a partir do interior, talvez o fizessem do exterior. Para isso recolheram alguns exemplares que estudaram no laboratório. O resultado desse estudo de Alexander Kupfer e colegas é descrito num artigo na Nature (ref2). Numa tradução livre do resumo:Apesar do crescimento inicial e desenvolvimento da maioria do animais multicelulares depender das provisões de vitelo, há muitas e variadas formas pelas quais os animais fornecem um investimento adicional ou alternativo aos seus descendentes.
O vitelo é a parte dos nutrientes num ovo, aquilo que vulgarmente se designa por gema.Fornecer nutrientes adicionais à descendência deveria ser favorecido pela seleção natural quando o aumento de aptidão dos jovens contrabalança qualquer redução correspondente na fecundidade. Formas alternativas de alimentação podem permitir aos progenitores retardar ou redirecionar o seu investimento. Relatamos aqui uma forma notável de cuidados parentais e o mecanismo de transferência de nutrientes entre progenitores e descendência num anfíbio ceciliano. Boulengerula taitanus é uma cecília ovípara de desenvolvimento directo, cuja pele é transformada nas fêmeas com crias por forma a providenciar uma fonte rica em nutrientes para a descendência em desenvolvimento. Esta forma de cuidados parentais fornece um estádio intermediário plausível na evolução da viviparidade nas cecílias. Aquando da independência, a descendência das cecílias dermatróficas vivíparas e ovíparas são relativamente grandes, apesar de lher ter sido fornecido pouco vitelo. A dentição especializada das cecílias que se alimentam da pele (dermatófagas) podem constituir uma pré-adaptação para a nutrição fetal nos tecidos do oviduto das cecílias vivíparas.
Para terminar nada melhor que mostrar alguma esfomeadas pequenas cecílias a alimentarem-se da pele da sua extremosa mãe.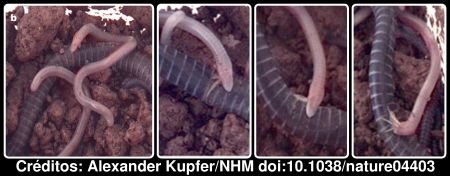
Referências
(ref1) Parker H.W. et al. (1956). VIVIPAROUS CAECILIANS AND AMPHIBIAN PHYLOGENY. Nature 178 250-252.
(ref2) Alexander Kupfer, Hendrik Müller, Marta M. Antoniazzi, Carlos Jared, Hartmut Greven, Ronald A. Nussbaum and Mark Wilkinson (2006). Parental investment by skin feeding in a caecilian amphibian. Nature 440, 926-929. Laço DOI.
quarta-feira, abril 12, 2006
Pinguins, imprensa e ADN com 60 milhões de anos
 Eu tento sempre acompanhar a descrição das descobertas, de que falo aqui no Cais de Gaia, de citações dos artigos originais, e em geral da tradução do resumo do artigo. Não se trata de uma questão de preguiça, mas apenas de mostrar o que vem na versão original, sem as especulações que acompanham a maioria dos comunicados de imprensa. Aliás, para lá das especulações, os meios de comunicação, ao adaptarem o formato, tamanho e conteúdo das notícias, para as tornarem ou mais curtas ou "mais interessantes", originam muitas vezes deturpações curiosas da mensagem original. Neste momento há milhões de pessoas que julgam que ADN de pinguins com mais de 60 milhões de anos está guardado algures num laboratório na Nova Zelândia. [... ler mais]
Eu tento sempre acompanhar a descrição das descobertas, de que falo aqui no Cais de Gaia, de citações dos artigos originais, e em geral da tradução do resumo do artigo. Não se trata de uma questão de preguiça, mas apenas de mostrar o que vem na versão original, sem as especulações que acompanham a maioria dos comunicados de imprensa. Aliás, para lá das especulações, os meios de comunicação, ao adaptarem o formato, tamanho e conteúdo das notícias, para as tornarem ou mais curtas ou "mais interessantes", originam muitas vezes deturpações curiosas da mensagem original. Neste momento há milhões de pessoas que julgam que ADN de pinguins com mais de 60 milhões de anos está guardado algures num laboratório na Nova Zelândia. [... ler mais]
Tudo começou com um artigo sobre a descoberta de um antepassado do pinguim moderno que teria vivido há mas de 60 milhões de anos, e do qual falámos aqui no dia 20 de Março. Ora a história foi acompanhada de um comunicado de imprensa, que foi adaptado pela Australian Broadcasting Company numa versão que não é clara acerca dos métodos usados para datar a descoberta nem acerca da forma como se estabeleceu que eram pinguins:New analysis of the world's oldest fossil penguins confirms some birds survived the mass extinction that killed the dinosaurs 65 million years ago, researchers say. The penguins once lived in shallow seas off New Zealand's east coast 60 million years ago. Now a molecular study, published in the journal Molecular Biology and Evolution, links them closely to modern penguins.
Uma tradução razoavelmente fiel do que está escrito acima seria:Uma nova análise dos fósseis mais antigos de pinguins confirma que algumas aves sobreviveram à extinção em massa que matou os dinossauros 65 milhões de anos atrás, dizem os investigadores. Os pinguins viveram em tempos em mares pouco profundos na costa este da Nova Zelândia 60 milhões de anos atrás. Agora um estudo molecular, publicado na revista Molecular Biology and Evolution, liga-os de perto aos pinguins modernos.
Neste trecho há logo algo que não está correcto. Determinou-se que os fósseis eram pinguins pelas propriedades do esqueleto, não por nenhuma análise molecular. Mas de resumo em resumo, a história foi encurtada um pouco mais e deu azo no Dominion Post ao que se transcreve abaixo.Dna tests on the Waimanu penguin fossils, found near the Waipara River, have determined they are between 60 million and 62 million years old - or up to 10 million years older than any other penguin remains discovered.
Que pode ser traduzido aproximadamente porTestes de ADN nos fósseis de pinguins do Waimanu, encontrados próximo do Rio Waipara, determinaram que têm entre 60 a 62 milhões de anos, ou mais 10 milhões de anos que quaisquer outros restos de pinguins encontrados.
Isto está completamente errado. Não foi assim que os fósseis foram datados, aliás nem se encontrou ADN nenhum nestes vestigíos. A idade dos fósseis foi determinada da forma tradicional, usando a estratigrafia do terreno, que por sua vez terá sido datada por comparação como outros terrenos, ou usando análises de isótopos radioactivos. O ADN degrada-se muito rapidamente e ao fim de uns poucos milhares de anos já há muito pouco com que trabalhar, mesmo em animais congelados ou usando partes mais robustas como a polpa dos dentes. Embora os métodos de sequenciação metagenómicos desenvolvidos recentemente tenham permitido sequenciar o ADN nuclear de mamutes com muitos milhares de anos, deve notar-se que nesse caso eram restos congelados e não fossilizados; essas técnicas nunca poderiam ser utilizadas em restos fossilizados com milhões de anos. Tal como discutimos aqui na contribuição do dia 20 de Março a análise de ADN que os cientistas fizeram foi a aves modernas ou extinctas recentemente (como as moas). A data de 60 milhões de anos do Waimanu foi usada pelos cientistas para calibrar os resultados, pois mostra que nessa altura os antepassados dos pinguins já se tinham diferenciado das restantes aves modernas.
Não deixa de ser interessante a forma como os meios de comunicação se limitam a processar informação sem verificar a fonte original. Como é bem sabido, cada vez que se reproduz algo com alterações de conteúdo a mensagem degrada-se e chega-se facilmente a algo muito diferente da mensagem original. Já agora, se alguém conseguir encontrar textos da cobertura jornalística deste achado em Portugal ou no Brasil, por favor enviem-me os laços. Gostaria de comparar.
Referências
(ref1) Kerryn E. Slack, Craig M. Jones, Tatsuro Ando, G. L. (Abby) Harrison, R. Ewan Fordyce, Ulfur Arnason, and David Penny (2006). Early Penguin Fossils, plus Mitochondrial Genomes, Calibrate Avian Evolution. Molecular Biology and Evolution. Laço DOI.
segunda-feira, abril 10, 2006
Prestes a sair da água

 Cientistas a trabalhar numa área remota do ártico descobriram os restos fossilizados de uma criatura, a que chamaram Tiktaalik roseae, que nos mostra que os peixes desenvolveram o equipamento que nos permite andar muito antes de terem invadido com sucesso terra firme. Já se sabia que os vertebrados terrestres descendiam de peixes pulmonados com barbatanas carnudas, que existiram há cerca de 385 milhões de anos, e cujos descendentes descobertos em rochas com 365 milhões de anos apresentam verdadeiros membros com dedos. Ora nesses 20 milhões de anos muito aconteceu, em particular a evolução das coisas que realmente fazem um tetrápode, como os pulsos, tornozelos, e dedos. [... ler mais]
Cientistas a trabalhar numa área remota do ártico descobriram os restos fossilizados de uma criatura, a que chamaram Tiktaalik roseae, que nos mostra que os peixes desenvolveram o equipamento que nos permite andar muito antes de terem invadido com sucesso terra firme. Já se sabia que os vertebrados terrestres descendiam de peixes pulmonados com barbatanas carnudas, que existiram há cerca de 385 milhões de anos, e cujos descendentes descobertos em rochas com 365 milhões de anos apresentam verdadeiros membros com dedos. Ora nesses 20 milhões de anos muito aconteceu, em particular a evolução das coisas que realmente fazem um tetrápode, como os pulsos, tornozelos, e dedos. [... ler mais]
O novo fóssil, Tiktaalik roseae, de aspecto vagamente crocodiliano, cai exactamente nesse hiato temporal, e mostra aquilo que seria esperado numa criatura intermédia entre um peixe completamente aquático e um vertebrado capaz de se deslocar em terra firme. A descoberta é descrita em pormenor em dois artigos na Nature. Numa tradução livre do resumo do primeiro (ref1) de Edward Daeschler e colegas:A relação entre os vertebrados com membros (tetrápodes) e os peixes com barbatanas lobadas (sarcopterígeos) encontra-se bem estabelecida, mas a origem das mais importantes características dos tetrápodes permanece obscurecida pela falta de fósseis que documentem a sequência de mudanças evolutivas. Relatamos aqui a descoberta de uma espécie fóssil bem preservada de um peixe sarcopterígeo do Devoniano Tardio do Ártico do Canadá que representa um imtermediário entre os peixes com barbatanas e os tetrápodes com membros, e fornece uma visão única de como e em que ordem importantes características dos tetrápodes apareceram.
Aqui convém abrir um parêntesis e dizer que os terrenos onde estes fósseis foram descobertos, hoje no círculo polar ártico do Canadá, na altura em que o Tiktaalik viveu se encontravam na proximidade do equador e gozavam de um clima de tipo tropical.Embora as escamas do corpo, os raios das barbatanas, a mandíbula inferior e o palato sejam comparáveis aos de sarcopterígeos mais primitivos, a nova espécie tem um topo do crânio mais curto, uma região dos ouvidos modificada, um pescoço móvel, uma articulação do pulso funcional, e outras características que prefiguram condições dos tetrápodes. As características morfológicas e o ambiente geológico deste novo animal são indicativos de uma vida em habitats maginais de águas pouco profundas.
Ora os três exemplares estavam tão bem preservados que um artigo inteiro é dedicado às barbatanas deste peixe desaparecido há cerca de 380 milhões de anos. As barbatanas peitoriais são descritas em grande detalhe num segundo artigo de Neil Shubin e colegas. Numa tradução livre do resumo:Pulsos, tornozelos e dedos distinguem os membros dos tetrápodes das barbatanas, mas evidência directa da origem dessas características não tem estado disponível. Descrevemos aqui os apêndices peitorais de um exemplar de um grupo irmão dos tetrápodes, Tiktaalik roseae, que são morfologicamente e funcionalmente transicionais entre uma barbatana e um membro.
 O uso do termo "grupo irmão" refere apenas o facto de que o Tiktaalik era apenas uma entre muitas espécies semelhantes e que é portanto pouco provável que seja um antepassado directo dos tetrápodes, mas seria muito possivelmente um parente próximo desse antepassado directo. Um pouco adiante:
O uso do termo "grupo irmão" refere apenas o facto de que o Tiktaalik era apenas uma entre muitas espécies semelhantes e que é portanto pouco provável que seja um antepassado directo dos tetrápodes, mas seria muito possivelmente um parente próximo desse antepassado directo. Um pouco adiante:A barbatana do Tiktaalik era capaz de uma vasto leque de posturas, incluindo uma com um membro apoiado no substrato na qual o ombro e o cotovelo estavam flectidos e o esqueleto distal estendido. A origem dos membros envolveu muito provavelmente a elaboração e proliferação de características já presentes nas barbatanas de peixes como o Tiktaalik.
De notar que embora os comunicados de imprensa estejam cheios de termos como "elo perdido" esse termo não é exacto neste caso. O caminho entre barbatanas e membros funcionais deve estar cheio de avanços e recuos. O que o Tiktaalik mostra de forma bastante clara é que os membros dos tetrápodes não são uma inovação assim tão grande, as barbatanas de peixes como o Titaalik já tinham a maioria dos ossos que mais tarde seriam importantes para construir membros capazes de andar em terra firme. Este fóssil é importante mas precisamos de muitos mais para podermos construir uma verdadeira árvore evolutiva. O que é óbvio para já é que a fronteira entre tetrápode e peixe não é assim tão abrupta como se poderia pensar. Para uma descrição muito detalhada desta descoberta, com uma elaborada refutação de alguma retórica religiosa criacionista aconselho a consulta do blog de Martin Brazeau, o Lancelet (em inglês).
Referências
(ref1) Edward B. Daeschler1 Neil H. Shubin and Farish A. Jenkins, Jr (2006). A Devonian tetrapod-like fish and the evolution of the tetrapod body plan. Nature 440, 757-763. Laço DOI.
(ref2) Neil H. Shubin, Edward B. Daeschler and Farish A. Jenkins, Jr (2006).The pectoral fin of Tiktaalik roseae and the origin of the tetrapod limb. Nature 440, 764-771. Laço DOI.
sexta-feira, abril 07, 2006
Mercenário Esloque
 Howard Tayler é um sujeito curioso. Com um bom emprego numa empresa de software nos Estados Unidos da América, casado, com filhos, decide um belo dia dedicar os seus a uma banda desenhada na Internet sobre um grupo de mercenários que navegam pela Galáxia em busca de serviços que exigem os seus talentos especiais. A arte ao início era, e estou a ser simpático, lamentável, mas com o tempo aprimorou-se, e hoje pode ser definida como "não é tão má como isso". Koalas genocidas, criaturas de matéria escura de Andrómeda empenhadas em destruir a Via Láctea, mercenários sem escrúpulos, viagens no tempo, clones, aqui há de tudo. Uma das coisas que sempre distinguiu esta banda desenhada, o Schlock Mercenary, de outras BDs na Internet foi que Howard Tayler cumpriu sempre escrupulosamente a actualização, nunca falhando com um dos painéis diários, nem mesmo o painel especial dos Domingos.
Howard Tayler é um sujeito curioso. Com um bom emprego numa empresa de software nos Estados Unidos da América, casado, com filhos, decide um belo dia dedicar os seus a uma banda desenhada na Internet sobre um grupo de mercenários que navegam pela Galáxia em busca de serviços que exigem os seus talentos especiais. A arte ao início era, e estou a ser simpático, lamentável, mas com o tempo aprimorou-se, e hoje pode ser definida como "não é tão má como isso". Koalas genocidas, criaturas de matéria escura de Andrómeda empenhadas em destruir a Via Láctea, mercenários sem escrúpulos, viagens no tempo, clones, aqui há de tudo. Uma das coisas que sempre distinguiu esta banda desenhada, o Schlock Mercenary, de outras BDs na Internet foi que Howard Tayler cumpriu sempre escrupulosamente a actualização, nunca falhando com um dos painéis diários, nem mesmo o painel especial dos Domingos.
Eu disse que se trata de um sujeito curioso porque às tantas decidiu abandonar o emprego bem remunerado e dedicar-se de corpo e alma ao Mercenário Esloque. Ele lá se tem aguentado, e acaba de lançar a primeira edição em papel da obra. A "edição de árvore morta" corresponde a uma aventura do meio da saga, e pode ser apreciada aqui das páginas 38 à 41 (pdf de qualidade média). Infelizmente existe apenas em Inglês. É claro que já encomendei a minha.
A marmota com cornos
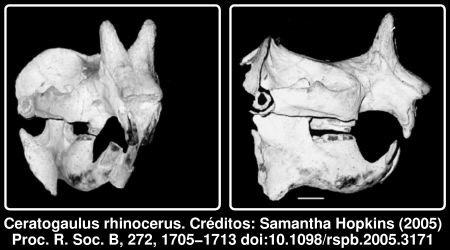
 Já que falei de roedores bizarros numa contribuição recente, não resisti hoje a desenterrar um artigo com um dos mais estranhos, e infelizmente extinto, membros do grupo. Não, não se trata de uma montagem, era realmente um roedor com dois cornos no focinho, o Ceratogaulus rhinocerus. Esta estranha criatura viveu nas planícies da América do Norte há alguns milhões de anos atrás, durante o Miocénico. O género Ceratogaulus inclui 4 espécies, que são os únicos roedores conhecidos com cornos. O facto de ser um animal fossorial, isto é, de hábitos subterrâneos, e com adaptações para escavar, torna a presença de chifres nasais, de tamanho respeitável nos últimos membros do grupo, ainda mais estranha. Os paleontólogos têm-se questionado sobre a função de tais estruturas, desde a sua descoberta, há mais de 100 anos. [... ler mais]
Já que falei de roedores bizarros numa contribuição recente, não resisti hoje a desenterrar um artigo com um dos mais estranhos, e infelizmente extinto, membros do grupo. Não, não se trata de uma montagem, era realmente um roedor com dois cornos no focinho, o Ceratogaulus rhinocerus. Esta estranha criatura viveu nas planícies da América do Norte há alguns milhões de anos atrás, durante o Miocénico. O género Ceratogaulus inclui 4 espécies, que são os únicos roedores conhecidos com cornos. O facto de ser um animal fossorial, isto é, de hábitos subterrâneos, e com adaptações para escavar, torna a presença de chifres nasais, de tamanho respeitável nos últimos membros do grupo, ainda mais estranha. Os paleontólogos têm-se questionado sobre a função de tais estruturas, desde a sua descoberta, há mais de 100 anos. [... ler mais]
É aqui que entra Samantha S. Hopkins, que estudou a história evolutiva e a morfologia destas estruturas. De acordo com os resultados dessa análise, publicada nos Proceedings of the Royal Society B (ref1), a melhor explicação para os chifres é que eles eram uma adaptação defensiva para evitar os predadores. Deve notar-se que o animal não era exactamente um peso-pluma, o traço branco na imagem acima corresponde a 1cm. Numa tradução livre do resumo:Ceratogaulus, um membro do clã extinto de roedores Mylagaulidae, é o único roedor conhecido com chifres, e o mais pequeno mamífero conhecido com chifres. A função deste grandes cornos nasais, que se projectam dorsalmente, num animal subterrâneo tem sido o sujeito de grande especulação entre paleontólogos; os usos sugeridos variam desde combate até escavação.
Aqui convém abrir um parêntesis e avançar um pouco no artigo, até onde a autora discute os vários tipos de fossorialidade nos roedores.Os roedores modernos usam um de três métodos para partir o solo. Tal como noutros mamíferos, a forma predominante de escavação entre os roedores é escavação por arranhar, no qual as garras das mãos são usadas para separar o solo. Muitos roedores também fazem uso dos seus incisivos, que crescem continuamente e que servem de diagnóstico ao clã, para escavar as suas tocas. Roedores que escavam fazendo uso dos incisivos superiores e inferiores, num movimento masticatório, são referidos como escavadores de dentes-de-escopro. Um último modo de escavação, e o menos comum entre os roedores, é a escavação por levantamento com a cabeça. Nos escavadores por levantamento da cabeça, o nariz, alguma vezes com a ajuda dos incisivos inferiores, é utilizado como uma pá para escavar a toca, usando os poderosos músculos do pescoço para levantar a cabeça.
Os ratos-toupeiros africanos, de que falámos aqui numa contribuição anterior, usam o método dos dentes de escopro. Ora qual era o método que os antepassados do Ceratogaulus utilizavam? Com base na morfologia das vértebras do pescoço, que sugerem músculos muito fortes, e no reforço da espessura dos ossos na zona do nariz, Samantha Hopkins conclui que o Ceratogaulus descende de roedores que escavavam por levantamento da cabeça. Sendo assim, é relativamente fácil ver como os chifres poderiam desenvolver-se numa criatura que já dispunha de um reforço na estrutura óssea na região do focinho. A questão é, poderiam esses chifres ser uma adaptação para facilitar o processo de escavação? A autora argumenta que não, muito pelo contrário.Os chifres nasais no Ceratogaulus são morfologicamente inconsistentes com o uso como ferramenta de escavação. Os chifres estão posicionados nas extremidades posteriores dos ossos nasais e estendem-se dorsalmente, perpendicularmente ao plano do palato. Em resultado da sua posição posterior, utilizar os chifres para escavar traria a extremidade anterior dos nasais contra o substrato após um muito curto movimento dos chifres, tornando a escavação com os chifres muito pouco eficiente.
Mais adiante nota ainda que a evolução vai no sentido oposto.Finalmente, os chifres posicionam-se mais atrás com o passar do tempo, pelo que a tendência evolutiva é pra um chifre que é adaptado de forma mais pobre para a escavação com o tempo. Logo, o argumento de que os chifres auxiliavam na escavação não é apoiado nem pela morfologia nem pela progressão evolutiva.
Ora se os chifres tornavam a vida fossorial mais difícil porque razão teriam aparecido? A autora avalia em seguida a questão de combate sexual, e afirma que é pouco provável porque os animais não mostram evidências de dimorfismo sexual, isto é, não se encontram evidências para diferenças entre os dois sexos. Todos os indivíduos apresentam em geral chifres semelhantes, e Samantha Hopkins nota ainda:Para além disso, muitas das objecções que se aplicam aos chifres como uma ferramenta de escavação também se aplicam ao uso dos cornos no combate sexual. A sua orientação, posição e a morfologia do resto do crânio tornam excessivamente difícil levá-los a encarar um oponente com um tamanho semelhante.
Para além disso, a autora nota que a visão destas criaturas era tão pobre que uma eventual fêmea a ver a competição nem se aperceberia de qual dos machos teria vencido o confronto, o que também é um argumento contra o uso dos chifres para que os indivíduos se reconhecessem entre si. Ora, para que surgiria então uma tal característica, com tão aparentes desvantagens para um criatura fossorial? Fosse qual fosse a razão teria que ser bastante importante, para se manter durante 10 milhões de anos, e para se tornar mais evidente com o tempo. A autora avança com uma explicação: defesa contra predadores. O Ceratogaulus era bastante maior que os seus predecessores, e teria que passar grande parte do seu tempo a alimentar-se à superfície num ambiente aberto (pradaria). Ora um animal pesado, de patas curtas, e grandes garras, seria presa fácil se não tivesse desenvolvido algum tipo de defesa. Samantha Hopkins avança então com o seguinte mecanismo, e analogia com um animal moderno.Os cornos são largos e robustos e a sua orientação dorsal e posição relativamente recuada tornam-nos adequados a protegerem os vulneráveis pescoço e olhos. Ao elevar a cabeça dorsalmente, os chifres seriam virados para trás, protegendo as áreas mais atacadas pelos predadores. Mostrou-se que uma utilização semelhante dos chifres posteriores-dorsais nos lagartos cornudos levou a um decréscimo da predação.
Um animal fascinante. Infelizmente não consegui encontrar uma ilustração do aspecto que teria em vida.
Referências
(ref1) Samantha S. Hopkins (2005). The evolution of fossoriality and the adaptive role of horns in the Mylagaulidae (Mammalia : Rodentia). Proceedings of The Royal Society B-Biological Sciences. 272 (1573), pp. 1705-1713. Laço DOI.
quinta-feira, abril 06, 2006
As formigas vertebradas

 Depois de ter falado em grilos que ocuparam o lugar dos ratos na Nova Zelândia, hoje foco o reverso da medalha. A criatura de hoje é um roedor que adoptou um estilo de vida característico dos insectos sociais como formigas e abelhas. O rato-toupeiro africano peludo, Cryptomys damarensis, vive em colónias de até 40 indivíduos, dominados por uma única fêmea reprodutora, e um ou dois machos seus parceiros. O resto da colónia descarrega as frustrações de uma vida de celibato numa única actividade: escavar com os seus grandes e poderosos incisivos, que se encontram no exterior da boca. Bem, na verdade, nem todos escavam. Boa parte da colónia, qualquer coisa como 40% dos indivíduos, passa a vida sem fazer grande coisa, e o papel desses "preguiçosos" confundia os cientistas. [... ler mais]
Depois de ter falado em grilos que ocuparam o lugar dos ratos na Nova Zelândia, hoje foco o reverso da medalha. A criatura de hoje é um roedor que adoptou um estilo de vida característico dos insectos sociais como formigas e abelhas. O rato-toupeiro africano peludo, Cryptomys damarensis, vive em colónias de até 40 indivíduos, dominados por uma única fêmea reprodutora, e um ou dois machos seus parceiros. O resto da colónia descarrega as frustrações de uma vida de celibato numa única actividade: escavar com os seus grandes e poderosos incisivos, que se encontram no exterior da boca. Bem, na verdade, nem todos escavam. Boa parte da colónia, qualquer coisa como 40% dos indivíduos, passa a vida sem fazer grande coisa, e o papel desses "preguiçosos" confundia os cientistas. [... ler mais]
Num artigo na Nature (ref1), Scantlebury e colegas mostram que nesses membros inactivos da sociedade dos ratos-toupeiros há mais um interessante paralelo com os insectos sociais. Numa tradução livre do resumo:A eussocialidade, que no mamíferos ocorre em apenas duas espécies de ratos-toupeiros africanos, caracteriza-se pela divisão de trabalho entre castas que se distinguem morfologicamente. Nos ratos toupeiros peludos (Cryptomys damarensis), o trabalho na colónia é dividido entre castas de "trabalhadores ocasionais" e "trabalhadores frequentes". Os trabalhadores frequentes estão activos durante todo o ano e em conjunto executam mais de 95% do trabalho total da colónia, enquanto os trabalhadores ocasionais executam em geral menos de 5% do trabalho total. Evidência de tipo anedótico sugeria que os trabalhadores ocasionais poderiam actuar como dispersadores, com a dispersão limitada ao comparativamente raros períodos em que o solo é amolecido pela água. Mostramos aqui que os trabalhadores ocasionais e as raínhas aumentam o seu dispender de energia após a queda de chuva, enquanto os trabalhadores frequentes não o fazem. Os trabalhadores ocasionais são também mais gordos que os trabalhadores frequentes. Sugerimos que os trabalhadores ocasionais constituem uma casta fisiologicamente distinta destinada à dispersão, cujos membros em vez de contribuirem para o trabalho de colónia e ajudarem a raínha a reproduzir-se, acumulam nos seus corpos reservas em preparação para a dispersão e reprodução quando as condições ambientais são as adequadas.
Ou seja, estes animais limitam-se a dormir e comer durante a maior parte do tempo, e quando desatam a escavar não é para ajudar ao trabalho na colónia, mas para a abandonarem. A vida fora da colónia é arriscada e até estabelecerem a sua própria colónia ou serem aceites noutra colónia as reservas de gordura fazem falta. Mas a raínha beneficia, pois são os seus genes que se vão espalhar.
O rato-toupeiro peludo não é o único mamífero eussocial, uma espécie aparentada, o rato-toupeiro africano nu, Heterocephalus glaber, ligeiramente menor, também vive em colónias que podem chegar aos 200 membros. Esta criatura é tão ou mais fascinante que o seu parente peludo, pois trata-se de uma espécie de mamífero ectotérmica, isto é, que não regula activamente a sua temperatura corporal. Além disso é seguramente um dos mais feios animais à face da Terra, e não resisto em mostrá-lo na imagem abaixo
Referências
(ref1) M. Scantlebury, J. R. Speakman, M. K. Oosthuizen, T. J. Roper and N. C. Bennett (2006). Energetics reveals physiologically distinct castes in a eusocial mammal. Nature 440, 795-797. Laço DOI.
domingo, abril 02, 2006
O parasita solitário que conduz a sua presa

 Imaginem uma criatura que devora a sua presa do interior e que irrompe de dentro dela para iniciar a sua vida como um predador destemido. Não, não se trata de nenhuma produção de Hollywood, a criatura existe mesmo, e este é apenas parte do tenebroso comportamento da pequena vespa iridiscente Ampulex compressa. Eu era para ter colocado isto ontem, mas depois pensei que os leitores iam olhar para a data e pensar que se tratava de uma partida do dia das mentiras. É mesmo verdade, e o mais arrepiante nesta história é que esta saída digna de um filme do Alien até nem é o aspecto mais perturbador. [... ler mais]
Imaginem uma criatura que devora a sua presa do interior e que irrompe de dentro dela para iniciar a sua vida como um predador destemido. Não, não se trata de nenhuma produção de Hollywood, a criatura existe mesmo, e este é apenas parte do tenebroso comportamento da pequena vespa iridiscente Ampulex compressa. Eu era para ter colocado isto ontem, mas depois pensei que os leitores iam olhar para a data e pensar que se tratava de uma partida do dia das mentiras. É mesmo verdade, e o mais arrepiante nesta história é que esta saída digna de um filme do Alien até nem é o aspecto mais perturbador. [... ler mais] Na verdade há algum tempo que eu queria falar desta criatura, e a a contribuição anterior serve como uma boa desculpa para falar aqui deste tema, que esteve em destaque no blog de Carl Zimmer durante o mês de Fevereiro. A pequena vespa iridiscente é um parasita durante a fase larvar que se especializou nesta espécie de baratas, e não ataca nenhumas outras. A Ampulex procura uma vítima e dá-lhe uma picada com o seu ferrão, injectando um cocktail de neurotoxinas que não mata a barata, apenas a paralisa ligeiramente. Este é apenas o início de uma história verdadeiramente tenebrosa. O comportamento da vespa é descrito em detalhe num artigo de Frederic Libersat no Journal of Comparative Physiology A (ref1). Numa adaptação livre do resumo:
Na verdade há algum tempo que eu queria falar desta criatura, e a a contribuição anterior serve como uma boa desculpa para falar aqui deste tema, que esteve em destaque no blog de Carl Zimmer durante o mês de Fevereiro. A pequena vespa iridiscente é um parasita durante a fase larvar que se especializou nesta espécie de baratas, e não ataca nenhumas outras. A Ampulex procura uma vítima e dá-lhe uma picada com o seu ferrão, injectando um cocktail de neurotoxinas que não mata a barata, apenas a paralisa ligeiramente. Este é apenas o início de uma história verdadeiramente tenebrosa. O comportamento da vespa é descrito em detalhe num artigo de Frederic Libersat no Journal of Comparative Physiology A (ref1). Numa adaptação livre do resumo:Porque o objectivo da vespa é fornecer uma refeição viva para a sua larva recém nascida, as mudanças de comportamento na presa são efectuadas pela manipulação do comportamento da presa numa forma que é benéfica para a vespa e para a sua descendência. Com esse fim, a vespa injecta o seu cocktail de veneno em duas ferroadas consecutivas. A primeira ferroada no tórax provoca uma paralisia temporária nas patas da frente que dura alguns minutos.
 A primeira ferroada é importante pois impede a barata de utilizar as patas da frente para afastar a vespa. É que o melhor está para vir. Este primeiro ataque químico é seguido por uma picada muito precisa na cabeça da infeliz barata, que injecta um cocktail de neurotoxinas directamente no cérebro da barata. O uso de neurotoxinas para incapacitar presas não é fora do comum. Cobras, escorpiões, aranhas, e mesmo caracóis utilizam-nos. Na maioria das vezes essas neurotoxinas perturbam o controlo das contrações dos músculos e levam à imobilização da presa. A ferroada da Ampulex faz algo mais, o cocktail de neurotoxinas injectado no cérebro da barata modifica o comportamento do insecto. A seguir à picada na cabeça, a barata desata a lavar as antenas e as patas da frente, durante cerca de meia-hora. Segue-se um outro padrão de comportamento que os autores designam por hipocinésia em que a barata fica indeferente a uma larga série de estímulos. O lavar e a hipocinésia só acontecem após a ferroada na cabeça. Se houver apenas a primeira ferroada no tórax, os efeitos passam ao fim de uns poucos minutos, e a presa recupera completamente.
A primeira ferroada é importante pois impede a barata de utilizar as patas da frente para afastar a vespa. É que o melhor está para vir. Este primeiro ataque químico é seguido por uma picada muito precisa na cabeça da infeliz barata, que injecta um cocktail de neurotoxinas directamente no cérebro da barata. O uso de neurotoxinas para incapacitar presas não é fora do comum. Cobras, escorpiões, aranhas, e mesmo caracóis utilizam-nos. Na maioria das vezes essas neurotoxinas perturbam o controlo das contrações dos músculos e levam à imobilização da presa. A ferroada da Ampulex faz algo mais, o cocktail de neurotoxinas injectado no cérebro da barata modifica o comportamento do insecto. A seguir à picada na cabeça, a barata desata a lavar as antenas e as patas da frente, durante cerca de meia-hora. Segue-se um outro padrão de comportamento que os autores designam por hipocinésia em que a barata fica indeferente a uma larga série de estímulos. O lavar e a hipocinésia só acontecem após a ferroada na cabeça. Se houver apenas a primeira ferroada no tórax, os efeitos passam ao fim de uns poucos minutos, e a presa recupera completamente. A hipótese do autor era que a vespa injectava o veneno directamente nos gânglios do cérebro e não apenas na vizinhança como alguns autores defendiam. Para isso colocou aminoacidos com C14 (radioactivo) nas glândulas de veneno das vespas e seguiu o trajecto do veneno nas baratas. Os resultados foram claros, a vespa injecta directamente em zonas precisas do cérebro. Para conseguir este tipo de precisão o autor especula que a vespa deverá ter orgão sensoriais no ferrão, embora não o verifique neste estudo. A fase de perda de sensibilidade a estímulos tem um objectivo simples.
A hipótese do autor era que a vespa injectava o veneno directamente nos gânglios do cérebro e não apenas na vizinhança como alguns autores defendiam. Para isso colocou aminoacidos com C14 (radioactivo) nas glândulas de veneno das vespas e seguiu o trajecto do veneno nas baratas. Os resultados foram claros, a vespa injecta directamente em zonas precisas do cérebro. Para conseguir este tipo de precisão o autor especula que a vespa deverá ter orgão sensoriais no ferrão, embora não o verifique neste estudo. A fase de perda de sensibilidade a estímulos tem um objectivo simples.Durante a hipocinésia de longa duração que se segue à lavagem, comportamentos específicos da presa são inibidos enquanto outros não são afectados. Propomos que o veneno reprime a actividade dos neurónios dos gânglios cerebrais removendo assim a pulsão excitatória aos neurónios do tórax.
O que se passa é que os mecanismos que levam à fuga das baratas aquando do ataque de um predador se localizam nos neurónios nos gânglios do tórax da barata. O que o autor propõe é que o efeito do veneno libertado no cérebro da barata leva à limitação das respostas desse circuito de fuga. Ora porquê tudo isto, por que é que a vespa não se limita a paralisar a barata?
A vespa precisa de levar a barata até ao seu ninho. Ora a vespa é demasiado pequena para arrastar a barata. Só que, devido a este elaborado esquema de ferroadas e toxinas, não precisa: basta-lhe agarrar numa das antenas da barata e passeá-la como um ser humano passearia um cão por uma trela. A barata limita-se a segui-la docilmente. Uma vez atingido o esconderijo a vespa deposita um ovo na barata, cobre o esconderijo com pequenas pedras, e vai-se embora, para não mais voltar. O trabalho com esta barata está feito e ela parte em busca de mais presas.
A larva desenvolve-se de início no exterior da presa, alimentando-se da hemolinfa da barata, mas ao fim de algum tempo entra dentro da barata onde se continua a alimentar, se transforma em pupa, e finalmente completa a metamorfose dando origem a mais uma pequena vespa iridiscente. A dramática sequência da eclosão da jovem vespa, seis semanas após a postura, pode ser apreciada no filme abaixo, cortesia de Fred Libersat, via Loom de Carl Zimmer.
Referências
(ref1) Fred Libersat (2003). Wasp uses venom cocktail to manipulate the behavior of its cockroach prey. J Comp Physiol A, 189: 497:508 Laço DOI.


