



 Este diabo da Tasmânia está simplesmente a descansar, mas o seu ar algo desanimado é adequado à situação periclitante destas criaturas. De seu nome científico Sarcophilus harrisii, o diabo da Tasmânia só se encontra nessa ilha ao largo da Austrália, tendo desaparecido do continente há cerca de 600 anos. Trata-se do maior carnívoro marsupial existente, do tamanho de um cão pequeno, medindo cerca de 60 centímetros de corpo-cabeça, com cerca de 25 centímetros de cauda. Pesa menos de 8 kg e, no estado selvagem, vive geralmente menos de 6 anos. Em 1996 foi detectado o primeiro caso de uma doença terrível, um tumor facial que acaba por levar à morte em poucos meses, e que tem dizimado populações. A doença aflige diabos em mais de metade da ilha e contina a espalhar-se. Por estranho que possa parecer, podemos estar a assitir a uma extinção causada por um animal morto há bastante tempo, mas que teima em assombrar os seus congéneres. [... ler mais]
Este diabo da Tasmânia está simplesmente a descansar, mas o seu ar algo desanimado é adequado à situação periclitante destas criaturas. De seu nome científico Sarcophilus harrisii, o diabo da Tasmânia só se encontra nessa ilha ao largo da Austrália, tendo desaparecido do continente há cerca de 600 anos. Trata-se do maior carnívoro marsupial existente, do tamanho de um cão pequeno, medindo cerca de 60 centímetros de corpo-cabeça, com cerca de 25 centímetros de cauda. Pesa menos de 8 kg e, no estado selvagem, vive geralmente menos de 6 anos. Em 1996 foi detectado o primeiro caso de uma doença terrível, um tumor facial que acaba por levar à morte em poucos meses, e que tem dizimado populações. A doença aflige diabos em mais de metade da ilha e contina a espalhar-se. Por estranho que possa parecer, podemos estar a assitir a uma extinção causada por um animal morto há bastante tempo, mas que teima em assombrar os seus congéneres. [... ler mais]
A explicação para a doença é da autoria de A.-M. Pearse e K. Swift e foi publicada na revista Nature (ref1) há alguns meses. Numa tradução livre do resumo:Uma parecença marcante no cariótipo destes tumores malignos significa que poderão ser infecciosos.
O diabo da Tasmânia, um carnívoro australiano de grandes dimensões, encontra-se sob a ameaça de doença fatal, amplamente espalhada, na qual um tumor da boca e da face perturba a capacidade do animal em se alimentar. Mostramos aqui que os cromossomas nestes tumores sofreram um rearranjo completo, que é idêntico para cada um dos animais estudados. À luz desta notável descoberta, e do bem conhecido comportamento agressivo dos diabos, propomos que a doença é transmitida por transplante alógeno, com uma estirpe de células infecciosas passada directamente entre os animais através de dentadas que infligem uns aos outros.
Os diabos da Tasmânia são animais solitários, mas que se juntam muitas vezes para se alimentarem de carcaças de animais mortos. Aí mostram maneiras à mesa execráveis, com rosnadelas, guinchos, e latidos constantes, e lutas terríveis em que se mordem uns aos outros, e os deixam cheios de cicatrizes. Como as dentadas ocorrem com maior frequência na região da boca, onde se localizam a maior parte dos tumores, os cientistas resolveram investigar se o tumor não estaria simplesmente a ser transplantado de uns animais para outros.
As células normais dos diabos possuem 14 cromossomas, que incluem os cromossomas sexuais XX e XY. As células cancerosas possuem apenas 13 cromossomas, que apresentam inúmeras anormalidades, e onde os cromossomas sexuais estão ausentes. O mais importante é que os tumores faciais em todos os animais investigados pelos autores apresentam exactamente as mesmas anomalias nas suas células cancerosas. A reorganização dos cromossomas dos diabos da Tasmânia é muito complexa, e não se detectaram estágios intermédios, mesmo em tumores pequenos que se estão a iniciar. Os autores referem que as modificações vistas nos cromossomas não são semelhantes a nenhum tipo de arranjo visto em tumores humanos, quer sejam espontâneos ou causados por vírus. Um dos animais estudados forneceu evidência bastante clara do que se está a passar:Apoio adicional, para a teoria do transplante alógeno na transmissão da doença, proveio da observação, afortunada, de uma inversão pericêntrica no cromossoma 5 no cariótipo de um animal. Esta anomalia foi encontrada em todas as culturas dos tecidos normais daquele diabo, mas não estava presente em nenhum dos cromossomas 5 das suas células do tumor facial, onde deveria ter sido observada se a neoplasia se originasse dos seus próprios tecidos.
A conclusão é simples: as células cancerosas não pertenciam originalmente a este animal, infectaram-no e desenvolveram-se no seu rosto. Os autores referem um tipo de tumor que mostra o mesmo tipo de anomalias nos cromossomas, o sarcoma venéreo transmissível dos cães, que se acredita ser também devido a um "clone capaz de existência parasítica." Pelo vistos há mesmo exemplos de transmissão de tumores cancerígenos em seres humanos:Os humanos podem também infectar-se uns aos outros acidentalmente com cancro (câncer), através da implantação de células em pacientes que receberam transplantes de orgãos. Esses cancros desenvolvem-se então de acordo com a sua progressão habitual.
Aqui os autores notam um aspecto importante para compreender a epidemia que afecta os diabos. Os transplantes de orgãos têm uma possibilidade de rejeição mais reduzida se o dador for um parente próximo, com tecidos compatíveis. Os diabos, com pouca diversidade genética, terão assim uma resposta imunitária reduzida às celulas cancerosas implantadas quando se mordem uns aos outros. Os seus sistemas imunitários não reagem a elas como sendo organismos invasores. Esta parece ser uma boa pista para compreender o que se está a passar no terreno. Para o passo seguinte vai ser preciso fazer análises de ADN mais detalhadas, que possam explicar a toxicologia, progressão, e propriedades epidemiológicas da doença.
O problema nesta história é que o tempo urge e as medidas têm que ser tomadas depressa. As taxas de mortalidade são aterradoras: são raros os animais que vivam mais de três anos, e a densidade populacional caiu qualquer coisa como 80% nos locais infectados. Para piorar as coisas, começaram a aparecer raposas na Tasmânia em 2001 e, se elas se estabelecerem, uma população reduzida e doente de diabos poderá não conseguir sobreviver.
Ficha técnica
Imagem do diabo a descansar cortesia de Wayne McLean, encontrada nesta página da Wikimedia Commons.
Referências
(ref1) A.-M. Pearse and K. Swift (2006). Allograft theory: Transmission of devil facial-tumour disease. Nature 439, 549. Laço DOI.
sexta-feira, outubro 20, 2006
Ter outro diabo no corpo
quinta-feira, outubro 19, 2006
O nobre e ameaçado rato-de-água

 O rato-de-água, da espécie Arvicola sapidus, é uma simpatica criaturinha, de pelagem espessa, que vive nas margens de cursos de água, canais de irrigação, lagos, e outras zonas húmidas, em Portugal, Espanha e França. Para roedor tem um tamanho razoável, uns respeitáveis 162 a 220 milímetros de cabeça-corpo, mais uma cauda com cerca de dois terços do comprimento do corpo. Pode atingir uns fabulosos 300 gramas de peso. Constrói galerias de túneis, em que uma das saídas é sempre submersa, e como o nome comum indica está muito bem adaptado à vida aquática. Essencialmente herbívoro, não desdenha insectos ou mesmo pequenos peixes, crustáceos, e anfíbios. Está em regressão, um eufemismo que se utiliza para dizer que caminha a passos largos para a extinção. Os motivos que me levam a falar neste roedor têm a ver com uma certa fanfarra na imprensa sobre um certo roedor de Chipre, que teria sido a única espécie de mamífero descrita na Europa nos últimos 100 anos. Pois bem o A. sapidus só ganhou esse nome em 1908. [... ler mais]
O rato-de-água, da espécie Arvicola sapidus, é uma simpatica criaturinha, de pelagem espessa, que vive nas margens de cursos de água, canais de irrigação, lagos, e outras zonas húmidas, em Portugal, Espanha e França. Para roedor tem um tamanho razoável, uns respeitáveis 162 a 220 milímetros de cabeça-corpo, mais uma cauda com cerca de dois terços do comprimento do corpo. Pode atingir uns fabulosos 300 gramas de peso. Constrói galerias de túneis, em que uma das saídas é sempre submersa, e como o nome comum indica está muito bem adaptado à vida aquática. Essencialmente herbívoro, não desdenha insectos ou mesmo pequenos peixes, crustáceos, e anfíbios. Está em regressão, um eufemismo que se utiliza para dizer que caminha a passos largos para a extinção. Os motivos que me levam a falar neste roedor têm a ver com uma certa fanfarra na imprensa sobre um certo roedor de Chipre, que teria sido a única espécie de mamífero descrita na Europa nos últimos 100 anos. Pois bem o A. sapidus só ganhou esse nome em 1908. [... ler mais]
Nas regiões tropicais descobrem-se constantemente novas espécies de vertebrados. Na Europa, com uma paisagem quase completamente talhada pelos humanos, e com uma enorme densidade de investigadores profissionais e naturalistas amadores, estas descobertas são mais raras. O exagero da imprensa é habitual, e neste caso inclui mesmo revistas com alguns pergaminhos. Por exemplo no comunicado de imprensa da National Geographic, é referido:A Europa ainda tem uns poucos segredos por descobrir. Cientistas anunciaram a descoberta do rato cipriota, a primeira nova espécie de mamífero lá descoberta em mais de um século.
Como mostra o A. sapidus é claramente menos de um século. De facto é bastante menos. Num comentário a esta notícia, Darren Naish, compilou uma lista de 30 espécies de mamíferos descobertas na Europa nos últimos 100 anos, no Tetrapod Zoology. A lista inclui roedores, musaranhos, toupeiras e morcegos, alguns deles descritos há muito pouco tempo. Há espécies de morcegos descritas já neste século. Mesmo de mamíferos terrestres há descobertas recentes, como um musaranho em 1989.
Embora o tal rato cipriota não seja "a descoberta do século", não deixa de ser importante e, quando o artigo científico for publicado, muito provavelmente colocarei algo aqui no Cais de Gaia. É pena que a excitação pela descoberta de uma nova espécie de roedor não seja de todo equivalente ao entusiasmo gerado pela conservação dos roedores já descritos. Se até lá a criaturinha não desaparecer, nas próximas férias vou fazer uma pequena expedição para tentar fotografar o rato-de-água lusitano, para substituir a imagem desta contribuição, para a qual não consegui indicação do fotógrafo nem de eventuais direitos de autor.
Ficha técnica
Comunicado de imprensa na National Geographic.
quarta-feira, outubro 18, 2006
Os orangotangos confusos



 Esta imagem de um orangotango, Pongo pygmaeus, escondido debaixo de uma saca, ilustra um pouco a nossa perspectiva relativamente a estes animais, pelo menos quando comparada com a visão que temos dos outros grandes símios. Quando se fala em animais não humanos inteligentes pensamos em chimpanzés, depois em gorilas, e raramente nos orangotangos. Trata-se de uma injustiça, pois, por exemplo, como vimos numa contribuição anterior, os orangotangos mostram um desempenho semelhante aos dos chimpanzés, gorilas, e crianças humanas de dois anos, nos chamados testes de deslocamento invisível. Resolvi por isso voltar ao tema das capacidades cognitivas dos grandes símios, embora sob uma perspectiva algo diferente, focando um teste em que todos falham. Curiosamente, os orangotangos, e todos os outros grandes símios, têm um desempenho muito pobre numa variante dos testes de deslocamento chamada tarefa de duplo deslocamento invisível. [.. ler mais]
Esta imagem de um orangotango, Pongo pygmaeus, escondido debaixo de uma saca, ilustra um pouco a nossa perspectiva relativamente a estes animais, pelo menos quando comparada com a visão que temos dos outros grandes símios. Quando se fala em animais não humanos inteligentes pensamos em chimpanzés, depois em gorilas, e raramente nos orangotangos. Trata-se de uma injustiça, pois, por exemplo, como vimos numa contribuição anterior, os orangotangos mostram um desempenho semelhante aos dos chimpanzés, gorilas, e crianças humanas de dois anos, nos chamados testes de deslocamento invisível. Resolvi por isso voltar ao tema das capacidades cognitivas dos grandes símios, embora sob uma perspectiva algo diferente, focando um teste em que todos falham. Curiosamente, os orangotangos, e todos os outros grandes símios, têm um desempenho muito pobre numa variante dos testes de deslocamento chamada tarefa de duplo deslocamento invisível. [.. ler mais]
Estava com uma certa preguiça em retomar este tema, e o regresso é motivado em parte por um certo desapontamento em não possuir a menor aptidão artística. Quando vejo, nesta página de pinturas premiadas de Carel Brest van Kempen, a terceira imagem, em que um orangotango olha com alguma curiosidade para os elefantes que se deslocam abaixo, sinto uma vontade imensa de ser capaz de criar algo semelhante. Infelizmente tenho que me ficar pela conversa, mas pelo menos é por uma boa causa. O artigo que fala do deslocamento duplo é da autoria de Joseph Call e foi publicado na revista Journal of Comparative Psychology (ref1). Numa tradução livre do resumo:Juvenis e adultos de orangotangos (n = 5; Pongo pygmaeus), chimpanzés (n = 7; Pan troglodytes), e crianças de 19 a 26 meses de idade (n = 24; Homo sapiens) foram confrontadas com deslocamentos visíveis e invisíveis. Foram apresentados três recipientes formando uma linha recta, e foi usada uma pequena caixa para deslocar uma recompensa debaixo delas. Os sujeitos foram confrontados com três tipos de deslocamento: simples (a caixa visitou um recipiente), duplo adjacente (a caixa visitou dois recipientes contíguos), e duplo não adjacente (a caixa visitou dois recipientes não contíguos). Todas as espécies tiveram desempenhos com níveis comparáveis, resolvendo todos os problemas excepto os deslocamentos invisíveis não adjacentes. Os deslocamentos visíveis eram mais simples que os invisíveis, e os deslocamentos simples eram mais simples que os duplos.
Os orangotangos têm um desempenho equivalente ao das outras espécies, inclusive nos testes em que falham. Não deixa de ser curioso que, quando se visitavam dois recipientes contíguos e depois se mostrava a pequena caixa vazia, os sujeitos da experiência não tenham grandes dúvidas em apontar um dos dos dois recipientes visitados. Mas, quando se visitavam os recipientes das extremidades saltando o do meio, muitas vezes as crianças e os antropóides escolhiam o do meio. O autor do estudo fez um outro teste onde este falhanço é mais óbvio:Numa segunda experiência, os sujeitos viram colocar as recompensas em dois recipientes adjacentes ou não adjacentes sem efectuar deslocamentos. Todas as espécies escolheram o recipiente vazio de forma mais frequente quando os recipientes com a recompensa eram não adjacentes do que quando eram adjacentes. Avançamos como hipótese que um enviesamento de resposta e um problema de inibição eram responsáveis pelo fraco desempenho em deslocamentos não adjacentes.
É sempre difícil interpretar resultados negativos. Será que o falhanço se deve aos atropóides e crianças de dois anos não possuirem as capacidades mentais necessárias, ou será que simplesmente acham a tarefa demasiado difícil e preferem não se dar ao trabalho? Os autores pensam que se trata provavelmente da segunda opção. Eu apresentei este artigo porque, tal como nos exemplos sobre a batota dos cães, mostra que é precisa muita cautela ao interpretar estudos sobre as capacidades cognitivas dos animais.
Eu referi acima que, quando se fala em animais não humanos inteligentes, se pensa em chimpanzés. Na verdade não é bem assim: para muitas pessoas o paradigma da inteligência animal são os golfinhos. As próximas contribuições sobre este tema irão focar os mamíferos aquáticos. Quão inteligentes são de facto os cetáceos?
Ficha técnica
Imagem da gorila no início da contribuição cortesia de Aaron Logan, retirada da sua galeria LIGHTmatter.
Referências
(ref1) Call J. (2001). Object permanence in orangutans (Pongo pygmaeus), chimpanzees (Pan troglodytes), and children (Homo sapiens). J Comp Psychol 115:159–171. Laço DOI.
terça-feira, outubro 17, 2006
Piolho de peixe


 Esta foto de Alexandra Morton mostra um parasita, o Lepeophtheirus salmonis, também chamado piolho do mar. Esta animal é um incómodo para os salmões adultos, embora em geral não coloque em grande perigo as suas vidas. As coisas são um pouco diferentes para os juvenis dos salmões, pois quando se tem menos de 3 centímetros de comprimento uma só destas criaturas pode ser suficiente para provocar a morte. Nas populações de salmão selvagem a mortalidade juvenil dos salmões devido a estes parasitas não é muito elevada. Isso sucede porque os piolhos do mar não conseguem sobreviver em água doce e assim os jovens salmões nascem num ambiente livre de piolhos. Quando os jovens seguem para o mar vivem durante bastante tempo longe dos cardumes dos adultos, pelo que a probabilidade de serem atacados pelos parasitas é muito pequena. O problema é que os seres humanos intrometeram-se nesta relação entre os salmões e os seus parasitas. [... ler mais]
Esta foto de Alexandra Morton mostra um parasita, o Lepeophtheirus salmonis, também chamado piolho do mar. Esta animal é um incómodo para os salmões adultos, embora em geral não coloque em grande perigo as suas vidas. As coisas são um pouco diferentes para os juvenis dos salmões, pois quando se tem menos de 3 centímetros de comprimento uma só destas criaturas pode ser suficiente para provocar a morte. Nas populações de salmão selvagem a mortalidade juvenil dos salmões devido a estes parasitas não é muito elevada. Isso sucede porque os piolhos do mar não conseguem sobreviver em água doce e assim os jovens salmões nascem num ambiente livre de piolhos. Quando os jovens seguem para o mar vivem durante bastante tempo longe dos cardumes dos adultos, pelo que a probabilidade de serem atacados pelos parasitas é muito pequena. O problema é que os seres humanos intrometeram-se nesta relação entre os salmões e os seus parasitas. [... ler mais]
As coisas mudaram com o aparecimento destas coisas que nos últimos decénios têm invadido zonas costeiras em muitas partes do mundo: Embora eu pessoalmente prefira sardinhas, e não goste por aí além de salmão, um grande número de pessoas parece considerar esse peixe uma iguaria, e as explorações de aquicultura intensiva de salmão têm crescido como cogumelos. Para além do carácter insensato desta actividade, pois trata-se um carnívoro que tem que ser alimentado com produtos de origem animal, a concentração de peixes num espaço tão pequeno levanta problemas ecológicos graves. Os salmões aprisionados num volume muito pequeno, com um volume de água por animal correspondente ao de uma banheira, contraem todo o tipo de doenças que transmitem às populações selvagens. Para além disso, os animais presos adquirem enormes cargas de parasitas. Durante algum tempo discutiu-se até que ponto o problema seria grave, com as multinacionais que controlam a aquicultura a tentarem pintar um quadro de tons suaves. Pois bem o debate acabou. O panorama é desastroso. Após um estudo exaustivo Martin Krkosek e colegas publicaram um artigo verdadeiramente arrepiante nos Proceedings of the National Academy of Sciences (ref1). Numa tradução livre do resumo:
Embora eu pessoalmente prefira sardinhas, e não goste por aí além de salmão, um grande número de pessoas parece considerar esse peixe uma iguaria, e as explorações de aquicultura intensiva de salmão têm crescido como cogumelos. Para além do carácter insensato desta actividade, pois trata-se um carnívoro que tem que ser alimentado com produtos de origem animal, a concentração de peixes num espaço tão pequeno levanta problemas ecológicos graves. Os salmões aprisionados num volume muito pequeno, com um volume de água por animal correspondente ao de uma banheira, contraem todo o tipo de doenças que transmitem às populações selvagens. Para além disso, os animais presos adquirem enormes cargas de parasitas. Durante algum tempo discutiu-se até que ponto o problema seria grave, com as multinacionais que controlam a aquicultura a tentarem pintar um quadro de tons suaves. Pois bem o debate acabou. O panorama é desastroso. Após um estudo exaustivo Martin Krkosek e colegas publicaram um artigo verdadeiramente arrepiante nos Proceedings of the National Academy of Sciences (ref1). Numa tradução livre do resumo:O declínio continuado das zonas de pesca e o aumento do consumo global de peixe levou a um aumento da aquicultura de 10% ao ano no último decénio. A associação das explorações piscícolas com o aparecimento de doenças em populações simpátricas de peixes selvagens permanece uma das ameaças que a aquicultura coloca aos ecossistemas costeiros e zonas pesca mais controversas e por resolver. Relatamos uma análise exaustiva da progressão e impacto de parasitas com origem em explorações piscícolas na sobrevivência de populações de peixes selvagens. Juntámos conjuntos de dados sobre a transmissão e patogeneicidade de piolhos do mar (Lepeophtheirus salmonis) em juvenis de salmões rosa (Oncorhynchus gorbuscha) e salmão chum (Oncorhynchus keta) migradores. Piolhos com origem nas explorações induziram uma mortalidade de 9-95% em várias populações simpátricas de salmões rosa e chum selvagens. As epizootias surgem através de um mecanismo que é novo para a nossa compreensão de doenças infecciosas emergentes: as explorações piscícolas sabotam o papel funcional da migração dos hospedeiros na protecção de hospedeiros juvenis de parasitas associados com hospedeiros adultos. Embora os ciclos de vida migratórios do salmão do Pacífico separem naturalmente os adultos dos juvenis, as explorações piscícolas fornecem ao Lepeophtheirus salmonis acesso a hospedeiros juvenis, neste caso aumentando as taxas de infecção durante pelo menos os primeiros 2.5 meses da vida marinha dos salmões (cerca de 80 km da rota migratória). A separação espacial entre juvenis e adultos é comum em peixes de águas temperadas, e com a continução do crescimento rápido da aquicultura, este mecanismo de doença pode desafiar a sobrevivência das economias e dos ecossistemas costeiros.
O ênfase é meu. Os pobres juvenis não têm hipótese, no seu caminho para o mar há uma nuvem de piolhos proveniente das explorações de salmão à qual não conseguem escapar. O salmão selvagem caminha para a extinção se a actividade das multinacionais de aquicultura intensiva continuar ou não for modificada. No Canadá, onde este estudo foi efectuado, não é apenas o salmão que está em perigo, todo o ecossistema fluvial que depende da migração dos salmões, incluindo por exemplo os ursos e águias pesqueiras, está em risco. Tudo para que alguns europeus e norte-americanos possam comer regularmente salmão. Como leitura de introdução ao tema recomendo a leitura do artigo da Watershed Watch Salmon Society (ref2).
Ficha técnica
Imensas imagens dos vários estágios de ataque por parasitas, da autoria de Alexandra Morton pode ser encontradas nesta página da Universidade Simon Fraser. Devo avisar que impressionam um bocado.
Referências
(ref1) Martin Krkosek, Mark A. Lewis, Alexandra Morton, L. Neil Frazer, and John P. Volpe (2006). Epizootics of wild fish induced by farm fish. Proc. Natl. Acad. Sci. USA, Laço DOI
(ref2) Watershed Watch Salmon Society (2004). Sea Lice and Salmon: Elevating the dialogue on the farmed-wild salmon story.PDF.
sexta-feira, outubro 13, 2006
Dia da família numerosa
 Todos os dias se celebra alguma coisa, nalgum lugar do mundo. Alguns países dedicam no entanto dias a coisas que à primeira vista causam estranheza. Os habitantes do Canadá escolheram uma família de insectos para "honrar" nesta data. Ainda por cima não se trata de insectos nobres como a abelha ou o bicho-da-seda: hoje no Canadá é o dia do gorgulho (caruncho). Isso mesmo, aqueles insectos "narigudos" que aparecem, volta e meia, no arroz ou cereais mal fechados e esquecidos durante algum tempo. Os gorgulhos partencem à família de insectos chamada Curculionidae que tem para cima de 40,000 espécies identificadas. Para situar bem este número, deve notar-se que foram descritas qualquer coisa como 5,500 espécies de mamíferos. Afinal talvez os gorgulhos mereçam algum reconhecimento. [... ler mais]
Todos os dias se celebra alguma coisa, nalgum lugar do mundo. Alguns países dedicam no entanto dias a coisas que à primeira vista causam estranheza. Os habitantes do Canadá escolheram uma família de insectos para "honrar" nesta data. Ainda por cima não se trata de insectos nobres como a abelha ou o bicho-da-seda: hoje no Canadá é o dia do gorgulho (caruncho). Isso mesmo, aqueles insectos "narigudos" que aparecem, volta e meia, no arroz ou cereais mal fechados e esquecidos durante algum tempo. Os gorgulhos partencem à família de insectos chamada Curculionidae que tem para cima de 40,000 espécies identificadas. Para situar bem este número, deve notar-se que foram descritas qualquer coisa como 5,500 espécies de mamíferos. Afinal talvez os gorgulhos mereçam algum reconhecimento. [... ler mais]
Dada a importância dos curculionídeos no que se refere ao número de espécies, resolvi por isso associar-me à data e colocar no Cais de Gaia esta imagem de dois Rhopalapion longirostre atarefados na tarefa que assegura a perpetuação da espécie.
Descobri o tal feriado do Canadá no blog Rigor Vitae: Life Unyielding, de Carel Brest van Kempen, que se define assim:Um biólogo falhado com habilitações para ser um varredor de ruas, porta-voz da NASA, ou artista da vida selvagem. Decidiu-se pela terceira opção.
Uma ilustração de um gorgulho, da sua autoria, pode ser vista na contribuição em que discute o tal dia do gorgulho no Canadá. Vale bem a pena uma visita. Bom dia do gorgulho para todos!
Já gora, em certas zonas de Portugal os gorgulhos também são chamados carunchos e não sei qual dos termos vingou no Brasil, ou se um outro termo qualquer é utilizado. A mão cheia de leitores brasileiros que passam por aqui, quase todos daquela que deve ser uma cidade magnífica, Curitiba no Paraná, que tenho que visitar um dia destes, talvez me possam informar.
Adenda. (16 Outubro 2006). Em mais um exemplo a juntar às muitas razões pelas quais não se deve acreditar em tudo o que se lê na internet, Carel Brest van Kempen confessa, nos comentários, que afinal o tal "feriado nacional" só é celebrado por uns quantos dos seus amigos.
Ficha técnica
Imagem de Rhopalapion longirostre cortesia de Jean-Jacques MILAN nesta página da Wikimedia Commons.
quinta-feira, outubro 12, 2006
Quando os camarões agitam as águas


 Esta imagem mostra o krill do Pacífico, de seu nome científico Euphausia pacifica, a sair do seu ovo e a iniciar o seu estádio larvar. Notem bem a escala: o traço corresponde a um décimo de milímetro. Ao completar o seu desenvolvimento esta espécie de krill cresce qualquer coisa como um factor de 100, atingindo um tamanho máximo que ronda os 2 centímetros, mas mesmo assim é uma criatura pequena. Só que pequeno é muito diferente de insignificante, muitas vezes as aparências enganam. Se, como indivíduos, não impressionam, as aglomerações destes camarões atingem números que desafiam a compreensão. Um trabalho recente, no fiorde de Saanich, na Colûmbia Britânica, estudou uma população onde havia qualquer coisa como 10,000 E. pacifica por metro cúbico, ou seja um animal a cada 5 centímetros. O mais incrível é que esse estudo sugere que este tipo de animais poderão desempenhar um papel importante, até agora ignorado, na produção de turbulência nos oceanos. [... ler mais]
Esta imagem mostra o krill do Pacífico, de seu nome científico Euphausia pacifica, a sair do seu ovo e a iniciar o seu estádio larvar. Notem bem a escala: o traço corresponde a um décimo de milímetro. Ao completar o seu desenvolvimento esta espécie de krill cresce qualquer coisa como um factor de 100, atingindo um tamanho máximo que ronda os 2 centímetros, mas mesmo assim é uma criatura pequena. Só que pequeno é muito diferente de insignificante, muitas vezes as aparências enganam. Se, como indivíduos, não impressionam, as aglomerações destes camarões atingem números que desafiam a compreensão. Um trabalho recente, no fiorde de Saanich, na Colûmbia Britânica, estudou uma população onde havia qualquer coisa como 10,000 E. pacifica por metro cúbico, ou seja um animal a cada 5 centímetros. O mais incrível é que esse estudo sugere que este tipo de animais poderão desempenhar um papel importante, até agora ignorado, na produção de turbulência nos oceanos. [... ler mais]
O estudo que mede a turbulência dos eufausídeos é da Eric Kunze e colegas e foi publicado na revista Science (ref1). Numa tradução livre do resumo:Medições numa região costeira mostraram turbulência que era três a quatro ordens de grandeza superior aquando da ascensão, durante o crepúsculo nocturno, de uma concentração de krill, aumentando a mistura diária no fiorde por um factor de 100.
Uma ordem de grandeza é a expressão científica para um factor de 10, um aumento de três a quatro ordens de grandeza significa 1,000 a 10,000 vezes mais. Mesmo tendo em linha de conta a curta duração da subida (e posterior descida) do krill isso é suficiente para que a mistura das águas nos vários níveis de profundidade seja 100 vezes maior que na ausência do krill. Este processo de mistura das águas tem influências óbvias na produtividade biológica local e até noutros temas muito em voga como o ciclo do carbono, e o clima:Porque as camadas de organismos nadadores que migram livremente se encontram em grande parte do oceano, turbulência gerada por processos biológicos pode afectar (i) o transporte de nutrientes inorgânicos para a superfície, muitas vezes pobre em nutrientes, a partir de níveis estratificados de águas subjacentes ricas em nutrientes, afectando a produtividade biológica, e (ii) a troca de gases atmosféricos como o CO2 com o interior do oceano estratificado, que não tem comunicação directa com a atmosfera.
Notem bem o ênfase no estratificado, termo que aparece várias vezes. É de facto importante notar que, contrariamente ao que possa parecer óbvio, a água no oceano mistura-se muito pouco entre as diferentes profundidades. Os processos de mistura são importantes pois controlam muitas das propriedades dos oceanos, em particular as trocas entre a camada mais superficial, em contacto com a atmosfera, e as regiões mais profundas. Os processos são muito lentos e são controlados pelo quebrar de ondas internas geradas pelos ventos e marés. Como esse ponto me parece importante, não resisto a citar partes do interior do estudo, que não fazem parte do resumo acessível a todos.As nossas medições tiveram lugar durante Abril de 2005. Durante esse período os ventos foram sempre muitos ligeiros. As águas abaixo de 5 metros eram fortemente estratificadas, com frequências de flutuação um centésimo de radiano por segundo. Estratificações ainda mais elevadas foram encontradas acima, com uma camada bem misturada com uma espessura não excedendo 3 metros.
Os autores seguiram as movimentações do krill usando métodos acústicos, ou seja ecos de ondas sonoras que permitem detectar a posição dos cardumes. Eco não é necessariamente o termo adequado mas confesso que não sei muito bem como traduzir backscatter. No que se segue vou utilizar retrodifusão, para me referir aos ecos enviados na direcção em que as ondas sonoras forma emitidas.Durante a luz do dia, o nível denso de krill retrodifusor permanecia estacionário a cerca de 100 metros, e as taxas de dissipação turbulentas eram próximas do nível de ruído instrumental de um milésimo de milionésimo de Watt por kg, um valor comparável aos sinais encontrados tipicamente em mar aberto.
Os 100 metros de profundidade não são um valor típico, mas têm aver com as características deste fiorde, os E. pacifica não podem descer mais pois as águas abaixo são anóxicas, devido à decomposição de matéria orgânica pelas bactérias. Os valores das taxas de dissipação dados aqui aqui são importantes sobretudo para comparação com o que vem a seguir. A mim dizem-me qualquer coisa, pois a turbulência é um tema em que estou particularmente à vontade, mas não se preocupem, não vou começar um tratado sobre o tema. O que é importante é ver como variam.No crepúsculo nocturno do dia 28 de Abril, a camada retrodifusora começou a subir e a ficar mais difusa. Inicialmente, não se observou nenhum aumento na microstrutura. Contudo, à medida que a base da camada retrodifusora (que se pensa estar associada aos eufausídeos maiores, com números de Reynolds suficentemente elevados para gerar turbulência) começou a migrar, os níveis de turbulência entre 30 a 100 metros de profundidade aproximaram-se de um décimo de milésimo a um centésimo de milésimo Watt por kg num intervalo de 10 a 15 minutos. Estes valores são 100 a 1000 vezes maiores que as taxas de dissipação no oceano estratificado profundo e são comparáveis aos valores encontrados em canais de maré turbulentos.
Aqui há vários pontos importantes. O krill migra em massa para a superfície durante a noite, com os mais pequenos a partirem uns minutos mais cedo que os maiores. São os maiores que geram a maior parte da turbulência. Notem a velocidade razoável, demoram 10 a 15 minutos a percorrer os 100 metros. Por outro lado passa-se de um milésimo de milionésimo a um centésimo de milésimo nos níveis de turbulência! Ou seja, de "zero ponto 8 zeros um" para "zero ponto 4 zeros um"; as tais quatro ordens de grandeza, ou seja um factor de 10,000 que referi atrás. Mesmo quando comparado com o que se passa no mar profundo o valor é 100 a 1000 vezes maior. O krill quando remexe as águas mistura-as de facto bem.
Os autores notam algo mais que também é interessante. As observações sugerem perturbações da ordem de 1 a 10 metros, o que não combina com os cerca de 1.5 centímetros dos indivíduos no cardume de krill. A explicação segundo eles é que os eufausídeos actuam de forma concertada, e não de forma individual, quando nadam para a superfície. Este comportamente é típico dos cardumes de muitas espécies, pelo que este processo não estará por isso limitado ao krill. Toda uma vasta gama de organismos grandes e pequenos, do zooplanctôn (0.5 cm) a cetáceos (da ordem de 10 metros) podem participar, e de facto gerar a maior parte dos níveis de turbulência nas camadas superiores do oceano, dominando os processos de mistura das águas nas regiões onde os organismos marinho são particularmente abundantes. Note-se em particular que isto é também é válido para o krill do Antárcticol E. superba, cujo papel importante no ciclo do carbono já tinha sido discutido aqui.
Nota: este artigo tinha-me passado um pouco "ao lado" quando li a Science. Só me dei conta dele quando li sobre ele no Deep-Sea News.
Ficha técnica
Imagem do krill a sair do ovo, cortesia de Jaime Gómez-Gutiérrez, obtida por intermédio da Wikimedia Commons, nesta página.
Imagem do cardume de krill obtida das páginas do Ocean Explorer da NOAA, aqui.
Referências
(ref1) Eric Kunze, John F. Dower, Ian Beveridge, Richard Dewey, and Kevin P. Bartlett (2006). Observations of Biologically Generated Turbulence in a Coastal Inlet. Science 313 (5794), 1768. Laço DOI
sexta-feira, outubro 06, 2006
O monstro com dentes grandes como pepinos

 Uma das muitas obras antigas, que encontrei no meu local de férias, foi o livro Water Reptiles of the Past and Present de Samuel Wendell Williston, publicado em 1914. Uma das figuras que ilustra esse livro é esta imagem de um Elasmosaurus platyurus, que se sempre me fascinou quando era miúdo. Quando se pensa em plesiossauros a imagem que nos ocorre é exactamente esta. Contudo, esta postura pescoço-de-cisne está errada: as vértebras articulavam de uma maneira que restringia muito a mobilidade do pescoço, em especial no que se refere aos movimentos verticais. Além disso, o peso do pescoço era bastante grande relativamente ao peso total do corpo pelo que o centro de massa teria que estar muito mais dentro de água. Mas o que me leva a falar em plesiossauros não é a postura do pescoço, mas sim uma descoberta recente:
Uma das muitas obras antigas, que encontrei no meu local de férias, foi o livro Water Reptiles of the Past and Present de Samuel Wendell Williston, publicado em 1914. Uma das figuras que ilustra esse livro é esta imagem de um Elasmosaurus platyurus, que se sempre me fascinou quando era miúdo. Quando se pensa em plesiossauros a imagem que nos ocorre é exactamente esta. Contudo, esta postura pescoço-de-cisne está errada: as vértebras articulavam de uma maneira que restringia muito a mobilidade do pescoço, em especial no que se refere aos movimentos verticais. Além disso, o peso do pescoço era bastante grande relativamente ao peso total do corpo pelo que o centro de massa teria que estar muito mais dentro de água. Mas o que me leva a falar em plesiossauros não é a postura do pescoço, mas sim uma descoberta recente:"Imaginem um animal tão comprido como um autocarro e com dentes tão grandes como pepinos. Ao mesmo tempo, imaginem que esse dentes se encontram numa cabeça na qual um homem adulto se pode deitar na língua e ser engolido inteiro."
[... ler mais]
Estas palavras não são minhas, mas de Jørn Hurum e Hans Arne Nakrem, que descobriram, na Noruega, um fóssil quase completo de plesiossauro que terá vivido há cerca de 150 milhões de anos. Este fóssil é parte de um achado mais vasto, foram identificados restos de 28 répteis marinhos. Dessa amostra 27 animais eram semelhantes aos que tinham sido identificados em 2004, numa expedição preliminar. O vigésimo oitavo, identificado a 5 de Agosto, era algo diferente e ultrapassou as expectativas. tratava-se do "monstro dos pepinos".
Um elasmossauro é na verdade tão grande como um autocarro, mas a menos que os pepinos na Noruega sejam minúsculos, os tais "dentes tão grandes como pepinos" não caberiam na sua boca. Ora o elasmossauro é um caso extremo, nem todos os plesiossauros tinham um pescoço tão grande como o resto do corpo. Alguns, como este Cryptoclidus, que media "apenas" quatro metros de comprimento, mostravam proporções menos exageradas.
Eis alguns exemplos, com vários níveis de "pescocidade", que podem ser encontrados nas páginas do Oceans of Kansas Paleontology de Mike Everhart:
Todos estes se enquadram, apesar de tudo, na ideia que a maior parte do público tem dos plesiossauros: algo que parece uma cobra enfiada dentro de uma tartaruga marinha. Há no entanto um grupo de plesiossauros que não se enquadram nessa imagem, os pliosauros, que se parecem com algo saído de um pesadelo. Estes eram poderosas máquinas assassinas, com bocarras enormes e aspecto feroz. O "monstro" da Noruega pertence a esse grupo. A que se assemelhava esse animal? Podemos ver aqui uma reconstituição do "monstro dos pepinos" a caçar um plessiosauro de menores dimensões. Estes animais apesar da "escassez" de pescoço são facilmente identificados como plesiossauros por possuirem 4 membros em forma de remos e caudas curtas. Uma criatura tão grande e armada de "pepinos" de tão grandes dimensões tinha pouco a temer das outras espécies de predadores no seu meio ambiente, e podia comer tudo o que conseguisse engolir, incluindo animais de uma outra família de répteis marinhos, os ictiossauros.
Estes animais apesar da "escassez" de pescoço são facilmente identificados como plesiossauros por possuirem 4 membros em forma de remos e caudas curtas. Uma criatura tão grande e armada de "pepinos" de tão grandes dimensões tinha pouco a temer das outras espécies de predadores no seu meio ambiente, e podia comer tudo o que conseguisse engolir, incluindo animais de uma outra família de répteis marinhos, os ictiossauros. Para aqueles que não ficaram grandemente impressionados, não resisto a mostrar mais imagens. Os pliossauros não são novidade, são conhecidos há muito tempo. O "monstro" é invulgar por ser um esqueleto quase completo. Para ver ilustrações de um outro pliossauro, provavelmente muito semelhante ao "dentes de pepino", sigam os apontadores abaixo para as páginas do Oceans of Kansas. Verifiquem pelo menos a primeira, adoro aquela imagem, parece realmente saída de um pesadelo dos mais aterradores.
Para aqueles que não ficaram grandemente impressionados, não resisto a mostrar mais imagens. Os pliossauros não são novidade, são conhecidos há muito tempo. O "monstro" é invulgar por ser um esqueleto quase completo. Para ver ilustrações de um outro pliossauro, provavelmente muito semelhante ao "dentes de pepino", sigam os apontadores abaixo para as páginas do Oceans of Kansas. Verifiquem pelo menos a primeira, adoro aquela imagem, parece realmente saída de um pesadelo dos mais aterradores.
Se estão com dificuldade em imaginar o tamanho destes bichinhos, nada melhor que imagens de humanos frente ao esqueleto de um Kronosaurus, que tinha um tamanho comparável ao do "monstro" norueguês e ao do Brachauchenius.
É quase impossível não ficar fascinado com tais criaturas. É claro que não podia falar numa descoberta paleontológica sem mostrar os fósseis. Eis aqui uma imagem das vértebras do "monstro dos pepinos" no local da escavação, com um dos descobridores, Jørn Hurum, a servir de escala. Nota: O comunicado de imprensa entretanto desapareceu pois houve progressos na história. Falo disso aqui.
Nota: O comunicado de imprensa entretanto desapareceu pois houve progressos na história. Falo disso aqui.
Ficha técnica
Não só imagens, mas também todo o tipo de informações sobre répteis marinhos extintos podem ser encontrados nas páginas de Mike Everhart, um dos melhores recursos de paleontologia na internet, o Oceans of Kansas Paleontology.
A foto do Cryptoclidus no Oxford University Museum of Natural History foi retirada desta página da Wikimedia Commons, user Ballista.
Referências
(ref1) Williston, S. W. 1914. Water Reptiles of the Past and Present. Chicago Univ. Press. 251 pp.
Macaquinho de imitação

 O macaquinho resus, de seu nome científico Macaca mulatta, não é tão desinteressante como o bocejo do ser humano deixa antever. Na verdade não se trata de um bocejo, este abrir da boca é parte de um trabalho científico bastante sério. Já referi aqui por diversas vezes que aquilo que os cientistas fazem nem sempre desperta grande entusiasmo e motivos de conversa na sua vida social. Por exemplo, como explicar que, depois de anos de estudo, faculdades, mestrados e doutoramentos, se ganhe a vida a fazer caretas a pequeninos macacos recém nascidos? Por estranho que pareça, os cientistas da imagem procuram resposta a algumas questões importantes, não estão apenas a tentar confundir o macaquinho. Neste caso trata-se de procurar compreender um comportamento que existe também nos bebés humanos. [... ler mais]
O macaquinho resus, de seu nome científico Macaca mulatta, não é tão desinteressante como o bocejo do ser humano deixa antever. Na verdade não se trata de um bocejo, este abrir da boca é parte de um trabalho científico bastante sério. Já referi aqui por diversas vezes que aquilo que os cientistas fazem nem sempre desperta grande entusiasmo e motivos de conversa na sua vida social. Por exemplo, como explicar que, depois de anos de estudo, faculdades, mestrados e doutoramentos, se ganhe a vida a fazer caretas a pequeninos macacos recém nascidos? Por estranho que pareça, os cientistas da imagem procuram resposta a algumas questões importantes, não estão apenas a tentar confundir o macaquinho. Neste caso trata-se de procurar compreender um comportamento que existe também nos bebés humanos. [... ler mais]
O pequenito da imagem acima, no seu primeiro dia de vida, face ao abrir da boca do ser humano, reagiu de forma semelhante. Mas nada melhor do que ver toda a cena em filme (AVI 3.3 Mbytes). Notem que o macaquinho dá sobretudo uma grande quantidade de "estalinhos" com os lábios.
Mas nada melhor do que ver toda a cena em filme (AVI 3.3 Mbytes). Notem que o macaquinho dá sobretudo uma grande quantidade de "estalinhos" com os lábios.
Esta capacidade dos macacos imitarem expressões não tinha ainda sido descrita na literatura. Pier Ferrari e colegas relatam-na pela primeira vez num artigo recente na revista PLoS Biology (ref1). Os autores estudaram 21 macaquinhos nos primeiro, sétimo, e décimo quarto, dias de vida. Numa tradução livre do resumo:A emergência de comportamentos sociais cedo na vida é provavelmente crucial para o desenvolvimento de relações mãe-filho. Pensava-se que alguns desse comportamentos, tais como a capacidade dos recém-nascidos imitarem movimentos faciais dos adultos, estariam limitados aos humanos e talvez à linhagem dos antropóides. Relatamos aqui as respostas comportamentais de macacos resus infantis (Macaca mulatta) às seguintes expressões faciais e gestos de humanos: estalar dos lábios, mostra da língua, abrir da boca, abrir da mão, e fechar e abrir dos olhos (condição de controlo). No terceiro dia de vida os macacos infantis imitam o estalar dos lábios e a mostra da língua.
Eis aqui o investigador a fazer o gesto ao macaquinho: A resposta não se fez esperar:
A resposta não se fez esperar: Mais uma vez a resposta pode ser seguida em filme (AVI 4.0 Mbyte). Não deixa de ser engraçado, ao contrário do que sucedia no filme anterior, notar a falta de interesse do macaquito pelo humano, pelo menos até ele começar a mostrar a língua. Mais uma vez o macaquinho dá imensos "estalinhos" com os lábios. Continuando com a introdução:
Mais uma vez a resposta pode ser seguida em filme (AVI 4.0 Mbyte). Não deixa de ser engraçado, ao contrário do que sucedia no filme anterior, notar a falta de interesse do macaquito pelo humano, pelo menos até ele começar a mostrar a língua. Mais uma vez o macaquinho dá imensos "estalinhos" com os lábios. Continuando com a introdução:No primeiro dia de vida, o abrir de boca do modelo suscitou um comportamento correspondente (estalar dos lábios) nos bebés. Estas respostas imitativas estão presentes num estádio inicial de desenvolvimento, mas estão confinadas aparentemente a uma janela temporal estreita. Como o estalar dos lábio é um gesto fulcral nas interações cara a cara dos macacos resus, a imitação neonatal pode servir para afinar as respostas de parentesco dos bebés ao seu mundo social. As nossas descobertas fornecem uma descrição quantitativa de imitação neonatal numa espécie primata não humana e sugerem que estas capacidades imitativas, ao contrário do que se pensava, não são exclusivas das linhagens humana e dos antropóides.Sugerimos que as suas origens evolutivas podem ser traçadas a gestos de filiação com funções comunicacionais.
Os autores referem que nos seres humanos há alguma variabilidade. Algumas crianças não mostram este tipo de comportamento, e apenas o deitar a língua de fora e abrir da boca têm um carácter mais ou menos geral. Nas crianças humanas este comportamento ocorre durante muito mais tempo que nos resus, durando cerca de 2 a 3 meses, período em as crianças aprendem a vocalizar e sorrir a outros seres humanos. Os chimpazés também imitam de forma semelhante a partir da primeira semana de vida até cerca dos 2 meses. Os autores referem no entanto que é preciso alguma cautela antes de considerar homólogas as respostas nos seres humanos e nos outros primatas. Há ainda muito poucos estudos feitos. Quanto às diferenças temporais do fenómeno entre os resus por um lado e chimpanzés e humanos por outro, os autores notam:Contrastando com os humanos e chimpanzés, os nossos bebés macacos mostraram o fenómeno durante apenas uns poucos dias após o nascimento. Tal como mencionado acima, alguns indivíduos ainda mostravam imitação do estalar dos lábios ao sétimo dias, mas não para além disso, Como se podem explicar tais diferenças entre espécies? O desenvolvimento motor e cognitivo é muito mais rápido nos macacos que nos humanos e chimpanzés. Logo à uma semana, os bebés macacos podem abandonar as suas mães durantes curtos períodos de tempo.
Embora seja preciso alguma cautela, pois os macaquitos estavam a interagir com seres de outra espécie, com os quais não tinham um relacionamento afectivo, parece tratar-se realmente de uma questão de precocidade no desenvolvimento:Nos humanos, e nos chimpanzés, os neonatos permanecem em contacto com as suas mães durante muito mais tempo, e a mãe é o único responsável por cuidar da criança. Logo, a imitação neonatal ocorre durante um período que, considerando os padrões típicos da espécie para o desenvolvimento de capcidades motoras e cognitivas, é comparável com que é descrito para humanos e chimpanzés.
Para além de terem mostrado que a noção de que os antropóides imitam, mas os macacos não, estava errada, os autores referem alguns resultados interessantes de outros trabalhos. Um que eu acho curioso é que os macacos percebem quando estão ser imitados pelos seres humanos.
Os macacos adultos também imitam prontamente os seus congéneres quando estes desempenham actividades relacionadas com a ingestão de alimento. Os macacos possuirão assim um mecanismo para ajustar o seu comportamento com o que observam nos outros indivíduos, embora em geral estejam limitados a um reportório de comportamentos típicos da sua espécie, que de certa forma estarão gravados nos seus cérebros. O comportamento é inato, não aprendido. Quanto ao mecanismo que explica este comportamento, os autores favorecem uma explicação baseada em descobertas neurofisiológicas, a dos espelhos neuronais, que se sabe existirem no cérebro dos macacos.Os neurónios nestes espelhos ficam activos quando o macaco executa uma acção específica com a sua mão (ou boca) e quando o macaco observa acções semelhantes com a mão (ou boca) executadas por outro indivíduo. Foi descrita recentemente uma classe de espelhos neuronais que responde a acções comunicadas por gestos faciais como o mostrar da língua ou o estalar dos lábios. Levando em consideração esses dados neurofisiológicos, os nossos dados concordam com a hipótese dos "espelhos neuronais", de acordo com a qual a observação de gestos com a boca activa directamente programas motores do mesmo tipo nas áreas pré-motoras do cérebro dos macacos, levando-as à ressonância e como consequência a darem lugar a uma réplica dos gestos observados.
Os autores referem ainda que a imitação neonatal nos humanos pode ser interpretada da mesma forma, e que há mesmo estudos que apoiam a existência de um sistema de espelho neuronal nos humanos homólogo aos dos macacos. Quanto à função deste comportamento, os autores sugerem que os macaquinhos estão a "afinar" os gestos que servem para identificar os indivíduos filiados num certo grupo. O comportamento de estalar os lábios, muitas vezes acompanhado de mostrar da língua é utilizado pelos macacos quando se aproximam para iniciar sessões de catamento. O artigo tem mais um vídeo interessante, de uma troca de estalidos de lábios entre mãe e filho (AVI 2.5 Mbytes) num cenário seminatural.
Não podia terminar sem mais imagens de macacos! Este artigo comprova, macaco vê: Macaco faz:
Macaco faz: Eu tinha já falado de macacos resus a propósito das vocalizações, neste artigo aqui. É sempre interessante descobrir mais pontos em comum com uma espácie cujo mais recente antepassado comum com os seres humanos terá vivido há mais de 25 milhões de anos.
Eu tinha já falado de macacos resus a propósito das vocalizações, neste artigo aqui. É sempre interessante descobrir mais pontos em comum com uma espácie cujo mais recente antepassado comum com os seres humanos terá vivido há mais de 25 milhões de anos.
Ficha técnica
Imagens e filmes retirados do artigo dado abaixo como ref1, e também da sinopse da autoria de Liza Gross, indicada como ref2
Referências
(ref1) Ferrari PF, Visalberghi E, Paukner A, Fogassi L, Ruggiero A, et al. (2006) Neonatal Imitation in Rhesus Macaques. PLoS Biol 4(9): e302. Laço DOI.
(ref2) Gross L (2006) Evolution of Neonatal Imitation. PLoS Biol 4(9): e311. Laço DOI.
quarta-feira, outubro 04, 2006
Apunhalar com preguiça

 Falei aqui recentemente daquele que teria sido o maior mamífero predador a caminhar sobre terra firme, com um peso que rondaria uma tonelada. Só que esse não era o único artigo sobre mega-carnívoros que encontrei entre a minha antiga literatura de férias. Alguns autores propuseram que o dono deste braço, o Megatherium americanum, seria também um predador. Este seria o cotovelo de um assassino. Ora o que é que isto tem de surpreendente? Bem, para já o megatério pesaria qualquer coisa como quatro toneladas, logo destronaria o Andrewsarchus por uma larga margem. Por outro lado, este braço pertence a nada mais nada menos que a uma preguiça gigante. [... ler mais]
Falei aqui recentemente daquele que teria sido o maior mamífero predador a caminhar sobre terra firme, com um peso que rondaria uma tonelada. Só que esse não era o único artigo sobre mega-carnívoros que encontrei entre a minha antiga literatura de férias. Alguns autores propuseram que o dono deste braço, o Megatherium americanum, seria também um predador. Este seria o cotovelo de um assassino. Ora o que é que isto tem de surpreendente? Bem, para já o megatério pesaria qualquer coisa como quatro toneladas, logo destronaria o Andrewsarchus por uma larga margem. Por outro lado, este braço pertence a nada mais nada menos que a uma preguiça gigante. [... ler mais]
Embora o artigo se mantenha algo controverso, devo confessar que, depois de falar de preguiças marinhas, nada me parece estranho nestes animais. Eis aqui uma versão de corpo inteiro deste alegado assassino.
O artigo é da autoria de dois cientistas uruguaios, R. Fariña e R. Blanco, e foi publicado na revista Proceedings of the Royal Society of London B (ref1). Numa tradução livre do resumo:O ponto de vista tradicional de que as preguiças terrícolas (Xenarthra) seriam um grupo relativamente uniforme e pouco diversificado do ponto de vista ecológico, foi desafiado recentemente. Hábitos marinhos foram atribuídos à Thalassocnus natans do Pliocénico do Peru. Além disso, uma dieta mais diversificada foi proposta por um de nós (R.A.F.) para alguns géneros de preguiças sul-americanas do Lujaniano (Pleistocénico tardio, Holoceno inicial). Neste artigo testa-se um aspecto da última hipótese, ou seja, que o Megatherium americanum tinha características morfológicas que se explicam melhor tendo hábitos carnívoros do que tendo uma dieta puramente herbívora. Abordamos, especificamente, a questão dos seus antebraços serem construídos por forma a optimizarem a velocidade em vez da força de extensão. Um tal traço poderia ter estado associado com um uso potencialmente agressivo das grandes garras do animal, enquanto uma extensão forte seria mais adequada para arrancar ramos. Por outro lado, a grande vantagem mecânica dos biceps poderia ter permitido ao animal levantar e carregar grandes pesos. Isto por sua vez sugere a possibilidade de que o animal poderia ter manipulado presas de grande tamanho (por exemplo, virando presas ou carcaças com armaduras dorsais para expor as parte mais desprotegidas) e para guardar grandes porções de comida em locais mais seguros. De acordo com esta visão, o Megatherium americanum seria o maior mamífero caçador terrestre a ter existido.
O que Fariña e Blanco mostram é que o olecrânio (porção do cúbito na vizinhança do cotovelo) dos carnívoros é optimizado para movimentos muito rápidos das antebraços. O megatério tem um olecrânio de carnívoro e os autores defendem que era utilizado pelo megatério para apunhalar outros animais. Os autores chegaram a essa conclusão depois de considerarem várias possibilidades, entre elas a defesa:Contudo, um mamífero adulto com um peso de 4 toneladas nos adultos não deveria estar particularmente preocupado com a possibilidade de ataque por, digamos um Smilodon dentes de sabre. Como analogia, um homem de 70 kg não deveria ter medo do ataque de um gato de 6 kg, independentemente da ferocidade que ele pudesse ter.
Eu confesso que o argumento não me convence totalmente. Eu teria medo se, em vez de um só bichano de 6 kg, fosse atacado por um bando deles com dentes que fizessem lembrar grandes adagas. Os autores consideram também não existir evidência de outras utilizações, como lutas entre indivíduos da mesma espécie. Daí pensarem que a única alternativa com sentido é o megatério ser carnívoro, embora não ser exactamente um assassino, talvez um oportunista que poderia roubasse as presas ao Smilodon. Contudo notam que o megatério podia adoptar uma postura bípede (ou trípode apoiado na cauda) e tinha membros anteriores muitíssimo fortes, sendo provavelmente capaz de levantar mais de uma tonelada. Isso seria útil para atacar gliptodontes. Estes eram grandes herbívoros couraçados, aparentados ao tatus, dos quais se mostra abaixo um exemplar da espécie Glyptodon asper. Os gliptodontes mediam cerca de quatro metros de comprimento e um metro e meio de altura, e pesariam por volta de uma tonelada e meia. É realmente uma pena que tenham desaparecido.
Os gliptodontes mediam cerca de quatro metros de comprimento e um metro e meio de altura, e pesariam por volta de uma tonelada e meia. É realmente uma pena que tenham desaparecido.
Como referi, a proposta de Fariña e Blanco foi acolhida com algum cepticismo. Em grande parte porque a dentição do megatério não é a de um carnívoro. As sínfises mandibular e maxilar do megatério não possuiam dentes, e os seus molares eram achatados. Não encontrei artigos recentes sobre o tema, pelo que a questão ainda estará por resolver, mas há para já uma imagem que não me sai da cabeça. Imaginem um bando de enormes preguiças assassinas a rodearem um pobre gliptodonte, a virarem-no de patas para o ar, e a apunhalarem-no até à morte com as suas enormes garras, furando-lhe a barriga desprotegida. Em seguida, imaginem essas criaturas, essencialmente desdentadas, excepto por um conjunto de molares nada adequados a mastigar carne, a retirarem sua vítima da carapaça pedaço por pedaço e a engolirem-lhe a carne. O mais interessante seriam as perseguições em câmara lenta entre as duas espécies, nenhuma delas realmente adaptada para correr depressa. Não sei, parece-me demasiado bizarro, mas na natureza já houve coisas mais estranhas.
Para aqueles com alguma curiosidade em ver o aspecto dos animais em vida, encontrei estas ilustrações de Heinrich Harder, da primeira metade do século XX, cujos direitos de autor expiraram e logo posso colocar aqui. Os seres humanos não estão a mais, foram de facto contemporâneos de ambas as criaturas nas Américas. Uma imagem mais dinâmica de um gliptodonte a ser atacado por uma matilha de canídeos pode ser vista nesta página de John Sibbick.
Os seres humanos não estão a mais, foram de facto contemporâneos de ambas as criaturas nas Américas. Uma imagem mais dinâmica de um gliptodonte a ser atacado por uma matilha de canídeos pode ser vista nesta página de John Sibbick.
Ficha técnica
Imagem do retirada desta página da Wikimedia Commons, user Ballista.
Imagem do gliptodonte retirada desta página da Wikimedia Commons, user Arent.
A arte de Heinrich Harder pode ser vista nesta página.
Referências
(ref1) Farina RA, Blanco RE. (1996). Megatherium, the stabber. Proc Biol Sci. 1996 Dec 22;263(1377):1725-9. Resumo e PDF
Quando a terra cospe lama

 Já falei aqui dos vulcões da Indonésia, dos seus jorros de lava, dos fluxo piroclásticos, das nuvens ardentes, e das colunas de cinza que projectam na atmosfera. Esta imagem ilustra um fenómeno que também está a ocorrer na Indonésia, e ao qual também é dado o nome de vulcão. Só que em vez de rocha fundida das entranhas da Terra, deste vulcão sai lama fervente. [... ler mais]
Já falei aqui dos vulcões da Indonésia, dos seus jorros de lava, dos fluxo piroclásticos, das nuvens ardentes, e das colunas de cinza que projectam na atmosfera. Esta imagem ilustra um fenómeno que também está a ocorrer na Indonésia, e ao qual também é dado o nome de vulcão. Só que em vez de rocha fundida das entranhas da Terra, deste vulcão sai lama fervente. [... ler mais]
Os vulcões de lama ou, mais propriamente, de materiais argilosos, formam-se quando material sedimentar, aprisionado com água suficiente para não cimentar por efeito da pressão, se acumula a profundidades de alguns km, na região de contacto entre placas tectónicas. Esses materiais podem subir ao longo de fracturas geológicas e atingir a superfície. Na maioria dos casos, esses vulcões de lama ocorrem no mar, e junto à costa portuguesa existem alguns, que têm sido estudados, não só pelo interesse científico, mas também porque estão muitas vezes associados a depósitos de hidrocarbonetos. Em terra firme são mais raros, existem por exemplo alguns em Timor, embora alguns dos mais espectaculares estejam no Azerbeijão. Por vezes ocorrem como resultado de perfurações levadas a cabo por companhias petrolíferas, mas expelem pouca lama e regra geral duram apenas alguns dias. Só que em geral, não quer dizer sempre, e este da Indonésia parece imparável.
O vulcão de lama indonésio originou uma imensa tragédia humana que prolonga há cerca de quatro meses na ilha de Java, próximo de uma localidade chamada Sidoarjo. Tudo começou a 29 de Maio, quando lama quente e vapores começaram a jorrar de um buraco próximo de um local onde estavam a ser executadas perfurações por uma empresa petrolífera. Não se sabe se a causa foram as perfurações ou tremores de terra que ocorreram dois dias antes. Desde então quatro povoações e 20 fábricas foram cobertas pela lama, que continua a fluir com valores que atingiram os 160,000 metros cúbicos por dia. A lama cobre já vários km quadrados com uma altura que excede 7 metros. Embora não haja registo de vítimas humanas, 11,000 pessoas perderam as suas casas. O fluxo de lama a ferver parece imparável, e segundo geólogos que visitaram o local é possível que a intervenção humana não consiga sustê-lo.
Neste momento a lama está a ser contida por uma rede de diques e pequenas barragens improvisadas, e com o começo da época das chuvas em Outubro o governo começou a permitir que alguma da lama seja largada num rio local chamado Porong que a transportará para o mar. Mas o problema é que isto não pode ser continuar por demasiado tempo. Um artigo da news@nature.com (ref1) cita o geológo Adriano Mazzini da Universidade de Oslo na Noruega:"Podem continuar a construir barragens, flitrar e despejar a lama eternamente. Supondo que ao fim de um ano não tenha parado, ter-se-á toda depositada no rio que terá que ser dragado."
De acordo com a news@nature.com, Mazzini sugere que se deva aproveitar a lama como matéria prima, fonte de energia geotérmica, ou para centros de férias para turistas interessados em banhos de lama. Claro que como geólogo Mazzini está sobretudo interessado nos aspectos científicos. Ainda uma citação do artigo da news@nature.com:"Estudos de um vulcão de lama que apareceu do nada nunca tinham sido feitos. Há todos os dados da companhia que fez as perfurações. É como se houvesse ali um laboratório."
O fenómeno que ocorre em Sidoarjo é um pouco mais complexo que um vulcão de lama clássico. Para além de ser gigantesco, a temperatura da lama faz pensar que se trata de uma espécie de híbrido entre um vulcão de lama clássico e uma fonte hidrotérmica como as que existem em Yellowstone.
Ficha técnica
Parte da inspiração para o texto e imagens retiradas desta página do grupo de Física de Processos Geofísicos (PGP) na Universidade de Oslo.
Referências
(ref1) Richard Van Noorden (2006). Java mud volcano seems unstoppable. Could Indonesia's mud flow be put to good use? news@nature.com. Laço DOI.
segunda-feira, outubro 02, 2006
Quando o magma sobe pelas fissuras

 Uma das coisas que me impressiona mais nas geociências é a capacidade de saber o que se encontra bem abaixo dos nossos pés, no interior da Terra. O vulcanismo é um dos fenómenos que nos permite esse tipo de conhecimento. Os materiais vulcânicos são uma das poucas fontes de informação directa sobre as condições existentes a grandes profundidades, em geral inacessível de outras formas. Isso sucede porque materiais arrastados ao longo do tubo de lava aparecem como inclusões nas rochas que se formam como resultado dos fenómenos vulcânicos. Essas inclusões vão desde fragmentos de rochas (xenólitos) a cristais indivíduais (xenocristais). Alguns exemplos de inclusões são bem visíveis na lava basáltica que se mostra na imagem, e que foi recolhida ao largo da Fossa do Japão. Ora este simples bocado de lava surprendeu os cientistas pois os dados de uma análise detalhada sugerem um mecanismo de formação de vulcões bastante diferente dos mecanismos que se admitiam até agora. [... ler mais]
Uma das coisas que me impressiona mais nas geociências é a capacidade de saber o que se encontra bem abaixo dos nossos pés, no interior da Terra. O vulcanismo é um dos fenómenos que nos permite esse tipo de conhecimento. Os materiais vulcânicos são uma das poucas fontes de informação directa sobre as condições existentes a grandes profundidades, em geral inacessível de outras formas. Isso sucede porque materiais arrastados ao longo do tubo de lava aparecem como inclusões nas rochas que se formam como resultado dos fenómenos vulcânicos. Essas inclusões vão desde fragmentos de rochas (xenólitos) a cristais indivíduais (xenocristais). Alguns exemplos de inclusões são bem visíveis na lava basáltica que se mostra na imagem, e que foi recolhida ao largo da Fossa do Japão. Ora este simples bocado de lava surprendeu os cientistas pois os dados de uma análise detalhada sugerem um mecanismo de formação de vulcões bastante diferente dos mecanismos que se admitiam até agora. [... ler mais]
Antes de passar ao estudo deste bocado de rocha tenho que abrir um pequeno parêntesis para introduzir o tema. Aqueles que leram a contribuição sobre a tomografia da região abaixo do Etna lembram-se que os mapas do que está abaixo da superfície eram feitos usando a velocidade das ondas sísmicas, que variam consoante o material atravessado, em particular se se trata de material sólido ou fundido. Foi o estudo dessas ondas sísmicas que permitiu identificar uma mudança nas propriedades de propagação alguns km abaixo da superfície, na transição da crosta para o manto. Esses estudos permitiram também descobrir que a camada do manto abaixo da crosta, até profundidades de muitas dezenas de km, é sólida e algo quebradiça, sujeita a fracturas. A crosta e esta camada do manto formam aquilo que se designa por litosfera, que cobre a superfície da Terra, e que está partida numa série de fragmentos, as placas tectónica. Abaixo da litosfera, existe uma zona, designada por astenosfera, onde o material tem uma consistência mais plástica, embora não esteja exactamente derretido, pois as ondas sísmicas de cisalhamento (ondas S) que não se propagam em fluídos propagam-se aí. A velocidade dessas ondas é contudo muito baixa, o que levou alguns autores a admitir que uma pequena fracção do material da astenosfera poderá de facto estar fundido, embora esta hipótese tenha sido questionada nalguns trabalhos recentes. Terminado o parêntesis posso então voltar ao fragmento de lava acima. A origem desse material são pequenos vulcões submarinos, com menos de 1km cúbico de material rochoso, as cercanias dos quais se mostram nas imagens abaixo, recolhidas por um veículo submersível. O que se está a ver são "almofadas de lava" com diâmetros de 30-50 cm, e pequenos calhaus resultantes do contacto explosivo de material aquecido com a água, de 4-6 cm de diâmetro, espalhados ao longo do cume do vulcão, que se encontra parcialmente coberto por sedimentos. Os cientistas ficaram um pouco surpreendidos quando observaram estes afloramentos rochosos pois não esperavam encontrar vulcões ali. O estudo detalhado destas rochas foi publicado recentemente na revista Nature (ref1) num artigo da autoria de Naoto Hirano e colegas. Numa tradução livre do resumo:
O que se está a ver são "almofadas de lava" com diâmetros de 30-50 cm, e pequenos calhaus resultantes do contacto explosivo de material aquecido com a água, de 4-6 cm de diâmetro, espalhados ao longo do cume do vulcão, que se encontra parcialmente coberto por sedimentos. Os cientistas ficaram um pouco surpreendidos quando observaram estes afloramentos rochosos pois não esperavam encontrar vulcões ali. O estudo detalhado destas rochas foi publicado recentemente na revista Nature (ref1) num artigo da autoria de Naoto Hirano e colegas. Numa tradução livre do resumo:Sabe-se que o vulcanismo na Terra ocorre em três cenários tectónicos: bordas de placas divergentes (tais como cordilheiras nos oceanos), bordas de placas convergentes (tais como arcos de ilhas), e pontos quentes.
A maioria dos vulcões ocorre na fronteira entre placas tectónicas. Os vulcões dos "pontos-quentes" que ocorrem no meio das placas tectónicas são mais raros. Segundo a hipótese que procura explicar esse fenómeno, em certos locais do manto, magma mais quente das profundezas seria, por acção de um mecanismo em grande parte desconhecido, empurrado num movimento ascendente, e como que fundiria a placa acima, sendo transportado até próximo da superfície. Como as placas tectónicas se deslocariam sobre essas "plumas do manto", razoavelmente fixas, o resultado desse movimento seriam cadeias de vulcões, como as do Havai. Os pequenos vulcões recém-descobertos localizam-se no meio de uma placa tectónica mas não se enquadram na categoria dos vulcões dos pontos quentes.Relatamos vulcanismo na Placa do Pacífico com 135 milhões de anos que não pertence a nenhuma dessas categorias. Pequenos vulcões alcalinos formam-se de pequenas percentagens de material fundido na astenosfera, como indicado pela sua geoquímica de elementos residuais e composição isotópica de gases nobres. Propomos que estes pequenos vulcões entra em erupção ao longo de fracturas na litosféricas em resposta à flexão da placa durante a subducção. Quantidades diminutas de fusão astenosférica e o alinhamento tectónico dos vulcões e a progressão em idade na direcção oposta ao do movimento da placa fornecem evidência para a presença de uma pequena percentagem de fusão na astenosfera.
O resumo é algo curto, e este artigo justifica que eu fale um pouco mais, em particular para que se possa compreender bem que os valores da idade e proveniência do material não surgem como que por magia. Os 135 milhões de anos são a idade da Placa do Pacífico na região da Fossa do Japão. A placa tectónica que os autores estudaram desloca-se qualquer coisa como 100 km a cada milhão de anos. Os pequenos vulcões são um fenómeno geológico bastante mais jovem que o fundo oceânico onde são transportados. Os mais próximos da Fossa do Japão, naquilo que os autores designam por local A, têm de 4.23± 0.19 a 8.5 ± 0.18 milhões de anos. A idade desses vulcões foi calculada através do estudo do decaimento de materiais radioactivos, usando o quociente entre o 40Ar e o 39Ar (Ar é o símbolo químico do árgon).
Os basaltos das "almofadas" recolhidas no local B, a cerca de 600 km do local A, possuem uma faixa de palagonite com uma espessura de 4 mm (está indicada na imagem no topo da contribuição). Os autores recorreram a essa faixa de palagonite para datar os basaltos, um método de datação que eu não conhecia. A lava submetida à acção da água sofre um arrefecimento brusco, formando um vidro que recobre a "almofada" de basalto. Ao longo do tempo, esse vidro sofre a acção hidratante da água, num processo de evolução estrutural e química que leva à formação de um mineral amarelado, a palagonite. A palagonite forma-se apenas dentro de água, e andou há uns tempos nas bocas do mundo por ter sido identificada em Marte, o que seria uma indício claro da existência de água no estado líquido. A palagonitização do vidro vulcânico exposto à água do mar não é rápida, pelo menos à escala humana, mas sim um processo contínuo de dissolução do vidro, formação de palagonite, e evolução dessa mesma palagonite. A taxa de crescimento é de ~0.003 a 0.02 mm por ano, logo 4 mm significam uma idade que pode ir de 50,000 anos a pouco mais de 1 milhão de anos para as lavas associadas. Ou seja, os vulcões mais afastados da Fossa do Japão são bastante mais jovens que os próximos da Fossa do Japão. A diferença de idade entre os dois locais é comparável ao tempo que a placa tectónica demora a percorrer os 600 km de distância entre os dois locais (cerca de 6 milhões de anos).
Segundo os autores do artigo os vários xenólitos e xenocristais incluídos nos basaltos fornecem alguns constrangimentos importantes quanto à origem das lavas:
Também segundo os autores, a lava mostra uma composição que sugere que se teria formado a profundidades superiores a 90 km. A Placa do Pacífico nesta região tem uma espessura de cerca de 95 km, o que indica que o magma provém de material fundido com origem na astenosfera. Por outro lado, os isótopos de gases raros presentes na lava mostram claramente que o material não teve origem nas camadas profundas do manto, como seria de esperar para o caso de vulcões de ponto quente, com origem numa pluma do manto. O esquema que os autores advogam é o seguinte: Os autores sugerem que a presença de materiais voláteis (essencialmente CO2) permite que exista uma pequena fracção de material fundido na astenosfera. Ainda segundo os autores, a subducção na Fossa do Japão leva a uma flexão da placa tectónica, que adopta uma forma convexa, com uma elevação de cerca de 800 metros acima do nível médio do fundo oceânico. A curvatura imposta na litosfera poderá então provocar algumas fracturas, permitindo que o material fundido da astenosfera suba e arraste os xenólitos litosféricos.
Os autores sugerem que a presença de materiais voláteis (essencialmente CO2) permite que exista uma pequena fracção de material fundido na astenosfera. Ainda segundo os autores, a subducção na Fossa do Japão leva a uma flexão da placa tectónica, que adopta uma forma convexa, com uma elevação de cerca de 800 metros acima do nível médio do fundo oceânico. A curvatura imposta na litosfera poderá então provocar algumas fracturas, permitindo que o material fundido da astenosfera suba e arraste os xenólitos litosféricos.
É óbvio que todos os comunicados de imprensa e mesmo a descrição na Science que acompanha este artigo, procuram um certo sensacionalismo, e falam logo em colocar em causa o mecanismo das plumas do manto. O artigo não vai tão longe, o que mostra mais uma vez o perigo de interpretar as descobertas científicas recorrendo a comunicados de imprensa. Os autores referem apenas que se trata de um outro mecanismo capaz de originar vulcões longe das bordas das placas tectónicas, e fornecem uma lista de outros locais semelhantes. Na verdade aquilo que os autores salientam com particular ênfase nas conclusões é a presença de material fundido na astenosfera, algo que tinha sido questionado recentemente.
Referências
(ref1) Naoto Hirano, Eiichi Takahashi, Junji Yamamoto, Natsue Abe, Stephanie P. Ingle, Ichiro Kaneoka, Takafumi Hirata, Jun-Ichi Kimura, Teruaki Ishii, Yujiro Ogawa, Shiki Machida, and Kiyoshi Suyehiro (2006). Volcanism in Response to Plate Flexure. Science Vol. 313. no. 5792, pp. 1426 - 1428. Laço DOI.
domingo, outubro 01, 2006
O cordeiro com pele de lobo

 Entre a literatura de anos anteriores, que se encontrava no meu local de férias habitual, encontrei um artigo publicado a 11 de Novembro de 1924, que descreve aquele que é o maior mamífero predador conhecido, o Andrewsarchus mongoliensis. Para aqueles que se interrogam sobre que idade terei, e se terei lido o tal artigo quando saiu, não, isso não aconteceu. Na verdade, embora tenha já um número apreciável de cabelos brancos, ainda não era nascido quando foi publicado. O animalzinho em questão, que viveu há cerca de 32 milhões de anos, era o dono do crânio que se mostra na figura, e na verdade pouco mais se conhece dele. Pode não parecer nada de especial, mas quando se refere o tamanho a coisa muda de figura: 834 milímetros de comprimento, apenas um pouco menos de metade da altura média de um homem adulto. [... ler mais]
Entre a literatura de anos anteriores, que se encontrava no meu local de férias habitual, encontrei um artigo publicado a 11 de Novembro de 1924, que descreve aquele que é o maior mamífero predador conhecido, o Andrewsarchus mongoliensis. Para aqueles que se interrogam sobre que idade terei, e se terei lido o tal artigo quando saiu, não, isso não aconteceu. Na verdade, embora tenha já um número apreciável de cabelos brancos, ainda não era nascido quando foi publicado. O animalzinho em questão, que viveu há cerca de 32 milhões de anos, era o dono do crânio que se mostra na figura, e na verdade pouco mais se conhece dele. Pode não parecer nada de especial, mas quando se refere o tamanho a coisa muda de figura: 834 milímetros de comprimento, apenas um pouco menos de metade da altura média de um homem adulto. [... ler mais]
A descoberta é descrita por Henry Fairfield Osborn na revista American Museum Novitates (ref1). O texto tem um certo sabor "histórico", por isso vou fazer uma citação bastante extensa. Numa tradução livre da introdução:O nome e descrição deste carnívoro omnívoro gigantesco, do Eocénico Superior da formação de Irdin Manha na Mongólia, são dedicados ao líder da Terceira Expedição Asiática, o senhor Roy Chapman Andrews. Quando primeiro descoberto pelo senhor George Olsen, foi declarado pelo senhor Andrews como um carnívoro, uma suposição que se mostrou ser correcta.
Roy Chapman Andrews é um personagem de certa forma mítico e atribuir o nome a um animal tão possante é um bom tributo. O artigo prossegue com:Posteriormente, o seu tamanho avantajado levou à suposição de que seria um membro da família de porcos gigantes representada pelo Entelodon na Europa e pelos Entelodontidae na América do Norte, - porcos omnívoros gigantes com crânios alongados; de facto, as proporções cranianas e faciais do Andrewsarchus são muitíssimo semelhantes às do Entelodon do Oligocene e do Dinohyus do Miocénico Inferior da América do Norte, sem dúvida devido a hábitos omnívoros semelhantes.
Uma visão artística de um entelodonte pode ser encontrada na imagem de marca que surge no alto de todas as páginas do blog de Carl Buel, o Olduvai George. O Entelodonte, que não é realmente um porco, tinha um crânio com mais de um metro de comprimento, e a sua dieta incluiria sobretudo plantas mas seria muito seguramente um necrófago oportunista e predador de pequenos animais. Rapidamente se verificou que o Andrewsarchus não era um entelodonte:Um rascunho dos contornos do crânio foi enviado numa carta ao Museu, a partir do qual o Doutor W. D. Matthew notou imediatamente a sua verdadeira afinidade como os primitivos creodontes da família Mesonychidae. Quando o exemplar chegou ao laboratório, foi comparado com o gigante Mesonyx (Harpagolestes) uintensis do Eocénico Superior do Wyoming.
A maioria dos mamíferos predadores a ques estamos habituados, como doninhas, lobos, ursos, hienas, felinos e pinípedes, pertencem a um grupo de animais chamados Carnivora. No início do século passado os creodontes eram um grupo que incluia practicamente todos os mamíferos placentários com habitos predatórios que não tivssem descendentes modernos. Esse não era um grupo natural, pois as relações de parentesco entre esses animais eram muitas vezes algo longínquas. Sabe-se hoje que muitos dos animais incluídos nos creodontes na altura em que o artigo Henry Osborn foi escrito estão de facto relacionados de mais perto com animais doutros grupos. Esse animais foram retirados dos creodontes, tendo os Creodonta sido redefinido como o "grupo-irmão" dos Carnivora. Os Carnivora e os Creodonta correspondem a dois ramos de mamíferos que possuem um antepassado comum mais recente entre eles do que com o resto dos mamíferos.
Nos últimos anos, com inúmeras novas descobertas, e a descrição de muitos fósseis guardados nas colecções dos museus, embora ainda haja muitas incertezas na filogenia dos mamíferos, sabe-se que os mesoniquídeos. que têm um registo fóssil bastante razoável, não são nem membros dos Carnivora nem dos Creodonta. Os mesoniquídeos, apesar de predadores, têm como parentes vivos mais próximos os artiodáctilos (um grupo que inclui entre outros ovelhas, camelos e hipopótamos) e os cetáceos. Daí o título desta contribuição: os mesoniquídeos eram parentes mais próximos dos cordeiros que dos lobos. Os mesoniquídeos são um exemplo interessante de evolução convergente com os canídeos se bem que com algumas peculiaridades. Possuíam uma forma vagamente canina, mas em vez de garras tinham um conjunto de pequenos cascos na ponta dos dedos. Uma reconstrução de um destes animais, do género Pachyaena, pode ser vista nesta página de Carl Buel, é a terceira figura do artigo. Trata-se de uma espécie bastante menor que o Andrewsarchus mongoliensis, mas o aspecto devia ser razoavelmente parecido.
Para perceber que o Andrewsarchus era mesmo grande basta ver o tamanho relativo do seu crânio comparativamente ao de outros carnívoros: três vezes maior que o do lobo e que o do seu parente próximo Mesonyx, e cerca de duas vezes maior que o do urso pardo.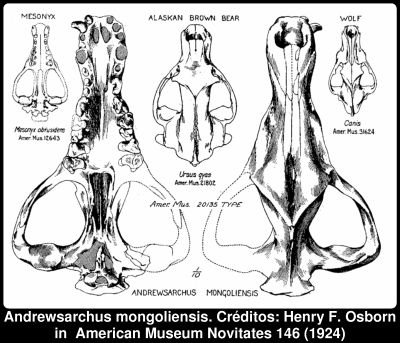 Mas exactamente quão grande era o Andrewsarchus? O problema é que só possuímos um crânio, logo temos que nos basear em estimativas obtidas a partir de parentes próximos para calcular as dimensões do resto do corpo. É isso que Henry Osborn faz para estimar o tamanho do animal:
Mas exactamente quão grande era o Andrewsarchus? O problema é que só possuímos um crânio, logo temos que nos basear em estimativas obtidas a partir de parentes próximos para calcular as dimensões do resto do corpo. É isso que Henry Osborn faz para estimar o tamanho do animal:Comprimento do corpo: tal como o comprimento do crânio de Mesonyx obtusidens (279 mm.) está para o comprimento total do corpo (1278 mm.), também está o comprimento do crânio do Andrewsarchus (834 mm.) para o seu comprimento total do corpo, nomeadamente 3820 mm.
Quase quatro metros de comprimento! Deve notar-se que o comprimento não inclui a cauda.Altura do corpo: tal como o comprimento do crânio de M. obtusidens (279 mm.) está para a altura das vértebras (576 mm.), também o comprimento do crânio do Andrewsarchus (834 mm.) está para a altura das vértebras, nomeadamente 1890 mm.
Quase dois metros de altura no garrote!Se o Andrewsarchus mongoliensis fosse proporcionado da mesma forma que o Mesonyx obtusidens, teria um comprimento do focinho até à extremidade do pélvis de 3 metros e 82 centímetros, e uma altura do chão ao garrote ou meio das costas de 1 metro e 89 centímetros. Assim, em números redondos, seria três vezes o tamanho do Dromocyon vorax do Mesonyx obtusidens do Eocénico Médio do Wyoming, formação de Bridger.
Quanto ao peso, Henry Osborn não avança estimativas. Mas não resisto a fazer uma estimativa pessoal baseada em animais vivos. Uma estimativa rápida e pouco precisa pode ser feita usando o factor de 3 no tamanho em relação aos lobos, a que deverá corresponder qualquer coisa como um factor de 27 no volume. Um lobo médio pesa menos de 60 kg, logo admitindo uma constituição semelhante o Andrewsarchus andaria por volta dos 1,600 kg. Esta forma de escalar os valores é um pouco simplista, mas, apesar disso, os valores que tenho visto na literatura, que usam aproximações mais adequadas, sugerem um animal apenas ligeiramente mais leve, com as estimativas variando entre os 900 e os 1,500 kg. Deve notar-se que temos apenas um exemplar, que não seria necessariamente o animal mais avantajado da espécie. De qualquer forma tudo sugere que existiu um mamífero predador pesando mais de uma tonelada e com uma bocarra com mais de 80 cm. Seria uma visão realmente assustadora.
Hoje em dia encara-se o Andrewsarchus como um predador possante, e não como um omnívoro. Uma visão artística de um Andrewsarchus mastigando um brontotério, enquanto uns creodontes do género Sarkastodon esperam pela sua vez, pode ser vista nesta página do artista John Sibbick.
Referências
(ref1) Henry Fairfield Osborn (1924). ANDREWSARCHUS, GIANT MESONYCHID OF MONGOLIA. American Museum Novitates, 146. PDF

