

 Eis que volto a deixar o meu blogue às moscas. Nesta imagem vêem-se dois machos da mosca-do-vinagre, Drosophila melanogaster, envolvidos numa cena de pancadaria. À margem da cena de pugilato, encontra-se uma terceira mosca, uma fêmea, caída por terra. A fêmea não adormeceu devido ao tédio causado pelo combate, o seu fim foi um pouco mais sinistro. Muitas vezes as fêmeas não se mostravam impressionadas pelos combatentes e iam-se embora. Depois de descobrirem que aos machos tanto fazia se a fêmea estava viva ou morta, os cientistas optaram por anestesiá-las com dióxido de carbono e decapitá-las antes de as colocarem nas arenas onde os machos se guerreavam. O trabalho por detrás desta cena macabra é muito sério: destina-se a estudar os fundamentos da agressividade, um dos aspectos mais importantes do comportamento animal. [... ler mais]
Eis que volto a deixar o meu blogue às moscas. Nesta imagem vêem-se dois machos da mosca-do-vinagre, Drosophila melanogaster, envolvidos numa cena de pancadaria. À margem da cena de pugilato, encontra-se uma terceira mosca, uma fêmea, caída por terra. A fêmea não adormeceu devido ao tédio causado pelo combate, o seu fim foi um pouco mais sinistro. Muitas vezes as fêmeas não se mostravam impressionadas pelos combatentes e iam-se embora. Depois de descobrirem que aos machos tanto fazia se a fêmea estava viva ou morta, os cientistas optaram por anestesiá-las com dióxido de carbono e decapitá-las antes de as colocarem nas arenas onde os machos se guerreavam. O trabalho por detrás desta cena macabra é muito sério: destina-se a estudar os fundamentos da agressividade, um dos aspectos mais importantes do comportamento animal. [... ler mais]
Por quê as moscas? Bem, para animais tão simples, com um cérebro menor que a cabeça de um alfinete, as moscas do vinagre possuem um comportamento elaborado. Os machos da espécie são territoriais e estabelecem relações de dominância. São também organismos baratos, que se podem produzir em grande quantidade, custam pouco a manter, e ocupam pouco espaço. Outra vantagem é que o genoma completo destes animais foi sequenciado, e pode ser manipulado para afectar de forma selectiva subconjuntos de neurónios do sistema nervoso central das moscas, o que permite identificar os mecanismos por detrás de traços do comportamento. Há ainda vantagens nos aspectos éticos: poucos grupos de activistas dos direitos dos animais se manifestam de forma tão violenta e irracional, quanto a experiências em moscas, como o fazem em relação a experiências em primatas ou ratos. Para além disso, há sempre supresas, mesmo em criaturas que se nos afiguram simples. É isso que sucede com o estudo referente à imagem: as moscas do vinagre possuem capacidades de memória e reconhecem mesmo outros indivíduos.
O estudo em questão, que envolve os aspectos da decapitação e lutas entre machos com intuitos necrofílicos, foi publicado por Alexandra Yurkovic e colegas na revista Proceedings of the National Academy of Sciences USA (ref1). Numa tradução livre do resumo:Os machos da Drosophila melanogaster (estirpe Canton-S) exibem agressividade na competição por recursos, ao defenderem o território, e para acesso a parceiros sexuais. No estudo descrito aqui, perguntámo-nos: (i) quanto tempo lutam as moscas; (ii) se as moscas adoptam estratégias distintas de vencedor e perdedor quando se estabelecem relações hierárquicas; (iii) se em lutas subsequentes, as moscas exibem mudanças na estratégia de luta decorrentes da experiência; e (iv) se as moscas lutam de maneira diferente em segundas lutas contra oponentes conhecidos ou desconhecidos.
Os pares de moscas macho lançadas na arena eram aquilo que os autores designam por animais ingénuos do ponto de vista social, isto é, que não tinham tido interações anteriores com indivíduos da mesma espécie. Dois dias antes da experiência as moscas macho foram anestesiadas com dióxido de carbono e foi-lhes feita uma marca com tinta acrílica que permitiu aos investigadores reconhecerem as moscas individualmente. Para além da "noiva-cadáver", também havia alimento em jogo: o copo onde as moscas lutavam possuía uma espécie de papa usada para alimentar as moscas, à qual tinham sido adicionadas umas gotinhas de sumo de maçã. Para terem um ideia da escala, o copo tinha um centímetro de altura, e um centímetro e meio de diãmetro. Trabalhar com estes animais tem realmente o seu quê de económico.
Continuando com o resumo:Os resultados mostraram que as moscas lutaram durante até 5 horas. À medida que se estabeleciam relações hierárquicas, as estratégias de comportamento mudavam: os vencedores progressivamente investiam mais e recuavam menos, enquanto os perdedores progressivamente investiam menos e recuavam mais. Os encontros entre as moscas eram frequentes durante os primeiros 10 minutos do emparelhamento mas depois caíam de forma significativa.
Ou seja, as moscas percebiam mais ou menos rapidamente qual seria o vencedor, que se mostrava cada vez mais agressivo, enquanto que o animal que percebia ser o perdedor, se mostrava mais receoso. Um aspecto pouco usual destes perdedores era que eles apesar de tudo enfrentavam repetidamente o vencedor. Os autores especulam que dado o facto de as moscas não possuirem armas de combate perigosas, não há um verdadeiro perigo à sobrevivência do perdedor, e na verdade é vantajoso para eles voltarem à superfície onde há alimento, embora mantenham as retiradas como o seu comportamento preferido. Os vencedores tendiam a permanecer na vizinhança da fêmea sem cabeça, onde passavam a maior parte do tempo, e o seu comportamento de combate preferido era investir na direcção de um perdedor que se aproximasse.Para responder a se as moscas recordavam combates anteriores, elas foram re-emparelhadas com opositores familiares ou não familiares após 30 minutos de separação. Nos emparelhamentos familiares, houve menos recontros durante os primeiros 10 minutos da luta que nos emparelhamentos com opositores não familiares, e antigos perdedores lutaram de forma diferente contra vencedores familiares que contra vencedores não familiares.
Este é um resultado surpreendente: as moscas do vinagre reconhecem indivíduos. Os autores referem que o mecanismo deste reconhecimento é desconhecido. Sugerem no entanto que durante os toques no corpo dos adversários durante os movimentos de investida, sejam capazes de se aperceber de diferenças nos hidrocarbonetos da cutícula. Um outro resultado inequívoco da experiência tem a ver com a memória que os perdedores guardam dos combates, que influencia o seu desempenho posterior:Os antigos perdedores perderam ou então não se chegou a uma decisão, em todos os segundos combates em emparelhamentos com vencedores conhecidos ou desconhecidos ou com moscas ingénuas. Emparelhamentos vencedor/vencedor, perdedor/perdedor, e ingénuo/ingénuo mostraram que os perdedores utilizaram estratégias de fraca intensidade em lutas posteriores e eram incapazes de formar novas relações hierárquicas, comparados com os vencedores ou com moscas socialmente ingénuas. Estes resultam apoiam fortemente a ideia de que a aprendizagem e memorização acompanham as mudanças de estatuto que resultam das lutas das moscas da fruta.
Pelos vistos os perdedores ficam "traumatizados" e tornam-se perdedores crónicos.
Ficha técnica
Imagem cortesia do Kravitz Lab da Harvard Medical School.
Referências
(ref1) Alexandra Yurkovic, Oulu Wang, Alo C. Basu, and Edward A. Kravitz (2006). Learning and memory associated with aggression in Drosophila melanogaster. PNAS 103: 17519-17524. Laço DOI.
quinta-feira, novembro 30, 2006
Os perdedores não esquecem
terça-feira, novembro 28, 2006
As taras da realeza

Pelo menos alguns patos têm vidas sexuais particularmente interessantes, envolvendo orgãos sexuais sobredotados, violações em bando e necrofilia ocasional. Algumas espécies são bizarramente agressivas, atacando e espancando outras aves aquáticas até à morte.
 Esta é uma citação de um texto de Darren Naish no Tetrapod Zoology sobre a redescoberta de uma espécie de pato que se julgava extinta. Depois de ter referido as moscas necrofílicas na minha última contribuição, a perversão entre os patos parece estranhamente adequada. O Darren foi atencioso e deu mesmo a referência. Foi daí que extraí esta imagem. Os animais em questão são dois patos-reais, da espécie Anas platyrhynchos, ambos machos. O que está caído no chão está morto, mas isso não deteve os avanços amorosos do segundo animal. [... ler mais]
Esta é uma citação de um texto de Darren Naish no Tetrapod Zoology sobre a redescoberta de uma espécie de pato que se julgava extinta. Depois de ter referido as moscas necrofílicas na minha última contribuição, a perversão entre os patos parece estranhamente adequada. O Darren foi atencioso e deu mesmo a referência. Foi daí que extraí esta imagem. Os animais em questão são dois patos-reais, da espécie Anas platyrhynchos, ambos machos. O que está caído no chão está morto, mas isso não deteve os avanços amorosos do segundo animal. [... ler mais]
Este comportamento é relatado por C. Moeliker, na revista Deinseaa (ref1). Numa tradução livre do resumo:A 5 de Junho de 1995 um pato-real macho adulto (Anas platyrhynchos) colidiu com a fachada em vidro do Natuurhistorisch Museum Rotterdam e morreu. Um outro pato da mesma espécie violou o cadáver quase continuamente durante 75 minutos. Então o autor perturbou a cena e recolheu o pato morto. A autópsia mostrou que a vítima de violação era de facto do sexo masculino. Conclui-se que os patos estavam envolvidos numa "Tentativa de Violação em Vôo" que resultou no primeiro caso de necrofilia homossexual nos patos-reais.
Eis aqui uma foto da parte mais perturbadora da história, tirada dois minutos após a que se mostra no início da contribuição. É de "admirar" a persistência do animal, 75 minutos, e mesmo assim ainda ficou algum tempo a protestar:
É de "admirar" a persistência do animal, 75 minutos, e mesmo assim ainda ficou algum tempo a protestar:Às 19:12 h eu perturbei esta cena cruel. O pato necrofílico deixou o seu "parceiro" de forma reluctante: quando eu me tinha aproximado dele a cerca de cinco metros, ele não levantou vôo mas andou simplesmente uns quantos metros, balbuciando fracamente séries de chamamentos de duas notas "raeb-raeb". Recolhi o pato morto e deixei o museu às 19:25 h. O pato ainda estava presente no local, chamando "raeb-raeb" e aparentemente procurando pela sua vítima (que, por essa altura, estava no frigorífico).
Embora tenha sido a colisão com o edifício que matou o pato, segundo o autor o animal devia estar a ser perseguido pelo outro que desejaria fazer dele, à força, o objecto das suas atenções amorosas. Aparentemente é um comportamento muito comum nos patos. Nunca mais vou encarar estes animais da mesma forma.
Referências
(ref1) C.W. Moeliker (2001). The first case of homosexual necrophilia in the mallard Anas platyrhynchos (Aves: Anatidae). Deinsea, vol 8, page 243-247. Resumo e PDF.
segunda-feira, novembro 27, 2006
Os combates das amazonas do vinagre, o musical
 Como resistir a estas esbeltas e longilíneas patas? À graciosidade destes tarsos delicados, terminados em elegantes garras? Aos sedosos e longos pêlos? Quem não se perderia no vermelho destes olhos, incandescentes como brasas? Não fora o mau feitio e esta Drosophila melanogaster seria a imagem da perfeição. Parece-me assim estranho que a minha incursão pela violência e pelo macabro na contribuição anterior, tendo como protagonistas as moscas do vinagre, não tenha surtido o efeito desejado. O número de visitas não disparou, muito pelo contrário. Ao procurar mais informação sobre os tais combates, encontrei aquela que me parece ser a explicação: falta a banda sonora. Essa é uma falha imperdoável, que tinha que corrigir. Aqueles com uma ligação de banda larga podem ver aqui o filme (26Mbytes) com acompanhamento musical do confronto entre fêmeas, cortesia do Kravitz Lab da Harvard Medical School.
Como resistir a estas esbeltas e longilíneas patas? À graciosidade destes tarsos delicados, terminados em elegantes garras? Aos sedosos e longos pêlos? Quem não se perderia no vermelho destes olhos, incandescentes como brasas? Não fora o mau feitio e esta Drosophila melanogaster seria a imagem da perfeição. Parece-me assim estranho que a minha incursão pela violência e pelo macabro na contribuição anterior, tendo como protagonistas as moscas do vinagre, não tenha surtido o efeito desejado. O número de visitas não disparou, muito pelo contrário. Ao procurar mais informação sobre os tais combates, encontrei aquela que me parece ser a explicação: falta a banda sonora. Essa é uma falha imperdoável, que tinha que corrigir. Aqueles com uma ligação de banda larga podem ver aqui o filme (26Mbytes) com acompanhamento musical do confronto entre fêmeas, cortesia do Kravitz Lab da Harvard Medical School.
Ficha técnica
Imagem de Drosophila melanogaster cortesia de André Karwath, retirada da Wikimedia Commons.
A necrofilia das moscas macho que lutam como meninas

 O número de leitores deste blog tem vindo a cair de forma acentuada, pelo que decidi tomar medidas drásticas antes que desça a zero. Resolvi assim apimentar um pouco as coisas, recorrendo às criaturas da imagem, as moscas do vinagre, Drosophila melanogaster. O macho (esquerda) e fêmea da imagem podem ter um aspecto simpático mas trata-se de animais aguerridos. Nesta contribuição vou falar de combates, trocas do comportamento típico dos sexos, e mesmo relações sexuais com cadáveres. Com filmes e tudo. Se isto não atrair mais visitas não sei o que o fará. [... ler mais]
O número de leitores deste blog tem vindo a cair de forma acentuada, pelo que decidi tomar medidas drásticas antes que desça a zero. Resolvi assim apimentar um pouco as coisas, recorrendo às criaturas da imagem, as moscas do vinagre, Drosophila melanogaster. O macho (esquerda) e fêmea da imagem podem ter um aspecto simpático mas trata-se de animais aguerridos. Nesta contribuição vou falar de combates, trocas do comportamento típico dos sexos, e mesmo relações sexuais com cadáveres. Com filmes e tudo. Se isto não atrair mais visitas não sei o que o fará. [... ler mais]
Eu tenho referido aqui por várias vezes que as actividades dos cientistas não dão lá grande tema de conversa em situações sociais. Encontrei mais um exemplo, num comunicado de imprensa relativo a um artigo sobre moscas do vinagre, de que falarei adiante. Nas palavras de Edward Kravitz, um estudioso da influência dos genes no comportamento animal:Quando se tenta explicar isso aos filhos -- "Pai, o que fizeste hoje?" "Bem, eu tinha estas duas moscas do vinagre, filho, e estava a ver se arranjava maneira de fazer com que lutassem." Pensem nisso.
Mas o mais delicioso é o que vem em seguida. Para levarem os machos a lutar os investigadores usavam a presença de fêmeas. Só que muitas vezes elas iam-se embora. A forma como os investigadores resolveram esse problema tem o seu quê de macabro. Ainda nas palavras de Kravitz:Quando transferiu a fêmea para o prato e acidentalmente lhe esmagou a cabeça, o meu estudante descobriu que os machos não se importavam se ela tinha cabeça ou não. Essa é a história verdadeira do que nos levou a cortar as cabeças das fêmeas nos estudos subsequentes.
Os machos realmente não queriam saber para nada das capacidades mentais das fêmeas:Eles cortejavam a fêmea morta, sem cabeça, e às vezes tentavam mesmo copular com ela.
Depois da parte com as referências à necrofilia vamos então aos combates. O estudo de Eleftheria Vrontou e colegas na revista nature neuroscience (ref1), foca o papel de um gene, que os autores chamam fruitless, a que eu me referirei adiante com infrutífero, no comportamento de combate das moscas do vinagre. Numa tradução livre do resumo:Quando competem por recursos, duas moscas Drosophila melanogaster do mesmo sexo lutam uma com a outra. Machos e fêmeas lutam com estilos marcadamente distintos, e os machos, mas não as fêmeas, estabelecem relações de dominância. Mostramos aqui que a formulação específica em função do sexo do gene infrutífero desempenha um papel fundamental no determinar com quem e como uma mosca luta, e se se forma uma relação de dominância.
O estudo é resultado de muitas horas de observação do comportamento das moscas do vinagre. Os autores arranjaram uma estranha arena, não percebo realmente do que se trata, que possuía alimento para atrair as moscas. Colocavam aí então um par de moscas e esperavam pelo confronto. Neste estudo não houve lugar a decapitações, nem necrofilia, isso foi apenas sensacionalismo barato para atrair leitores (não é totalmente verdade, irei falar desse estudo no futuro).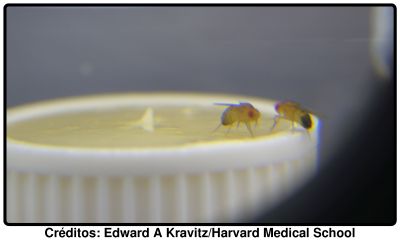 Em condições normais, as moscas macho, quando competem por algo, lutam de uma forma em que vale tudo: empurrões, cabeçadas e mesmo o equivalente drosofiliano de murros. Um filme (185 kbytes) da coisa acompanha o artigo. É verdadeiramente esclarecedor, trata-se de tareia a sério. As fêmeas lutam de forma mais delicada, tal como se pode ver neste outro filme (267 kbytes). Tudo muito suavezinho: uns empurrões ligeiros aqui, uns toquezitos com a cabeça ali. Enfim, coisa de meninas, só faltou puxarem as antenas uma à outra.
Em condições normais, as moscas macho, quando competem por algo, lutam de uma forma em que vale tudo: empurrões, cabeçadas e mesmo o equivalente drosofiliano de murros. Um filme (185 kbytes) da coisa acompanha o artigo. É verdadeiramente esclarecedor, trata-se de tareia a sério. As fêmeas lutam de forma mais delicada, tal como se pode ver neste outro filme (267 kbytes). Tudo muito suavezinho: uns empurrões ligeiros aqui, uns toquezitos com a cabeça ali. Enfim, coisa de meninas, só faltou puxarem as antenas uma à outra.
Ora machos e fêmeas, em condições normais, possuem formas distintas do tal gene infrutífero. Quando os investigadores colocaram o gene típico das fêmeas nos machos eles passaram a lutar "como meninas", e vice-versa. Vejam aqui o desempenho de combate de dois machos com o géne infrutífero fêmea em filme (269 kbytes). Lutam realmente como meninas.
Este estudo não se destina apenas a ver luta livre de moscas, por muito interessante que ela possa parecer. O objectivo é muito sério. Trata-se de compreender como funciona a agressividade. Pelos vistos nas moscas um só gene tem um papel fulcral na forma como o sistema nervoso controla esse comportamento. O próximo passo é obviamente passar da genética para a neurociência, isto é, compreender o que se passa nos neurónios das mosquinhas.
Confesso que fiquei fascinado com as cenas de pugilismo entre as moscas do vinagre. Ainda andei ver na minha cozinha mas não encontrei nenhuma, e com o frio que faz por aqui não vale a pena deixar a janela aberta para ver se atraio alguma. Já agora, não é a primeira vez que falo aqui de drosófilas. Já tinha falado dos gigantescos espermatozóides novelo da D. bifurca nesta contribuição.
Ficha técnica
Comunicado de imprensa de onde foram retiradas as citações iniciais pode ser encontrado aqui.
Imagens das moscas do vinagre cortesia da NASA (é isso mesmo estes insectos foram ao espaço).
Referências
(ref1) Eleftheria Vrontou, Steven P Nilsen, Ebru Demir, Edward A Kravitz & Barry J Dickson (2006). fruitless regulates aggression and dominance in Drosophila. Nature Neuroscience vol 9, 1469-1471. Laço DOI.
domingo, novembro 26, 2006
Paleogenómica humana

 Vivemos neste momento uma verdadeira revolução na biologia com o aparecimento da genómica, em particular com o conhecimento dos genomas completos dos seres vivos. Depois da fanfarra que acompanhou o anúncio da sequenciação do genoma humano, outros se têm seguido a um ritmo razoável. Nos tempos mais recentes lembro-me do genoma do chimpanzé, da abelha melífera, e do ouriço do mar. São tempos fantásticos de descoberta, e isto é apenas o prenúncio do que está para vir. Mas se a genómica é já de si algo fantástico, nos últimos tempos tem-se falado com grande insistência numa coisa chamada paleogenómica, ou seja a sequenciação do genoma de criaturas extintas. Entre essas criaturas extintas a serem estudadas inclui-se mesmo um membro do género Homo, nada mais nada menos que o Homo neanderthalensis. [... ler mais]
Vivemos neste momento uma verdadeira revolução na biologia com o aparecimento da genómica, em particular com o conhecimento dos genomas completos dos seres vivos. Depois da fanfarra que acompanhou o anúncio da sequenciação do genoma humano, outros se têm seguido a um ritmo razoável. Nos tempos mais recentes lembro-me do genoma do chimpanzé, da abelha melífera, e do ouriço do mar. São tempos fantásticos de descoberta, e isto é apenas o prenúncio do que está para vir. Mas se a genómica é já de si algo fantástico, nos últimos tempos tem-se falado com grande insistência numa coisa chamada paleogenómica, ou seja a sequenciação do genoma de criaturas extintas. Entre essas criaturas extintas a serem estudadas inclui-se mesmo um membro do género Homo, nada mais nada menos que o Homo neanderthalensis. [... ler mais]
Nas últimas semanas foram divulgados dois artigos sobre a sequenciação de partes significativas do genoma do homem de neandertal. Eu tinha referido os trabalhos em curso numa contribuição anterior, mas desta vez é mesmo a valer. Ambos os trabalhos usam um mesmo fragmento de um neandertal encontrado numa gruta na Croácia e que teria vivido há qualquer coisa como 38,000 anos. Um dos estudos recuperou já mais de 1 milhão de pares de bases (os constituintes do ADN) de material genético desse neandertal. Esse artigo, de Richard Green e colegas, foi publicado na revista Nature (ref1). Numa tradução livre do resumo:Os neandertais são o grupo hominídio extinto que está relacionado de mais perto com os humanos contemporâneos, pelo que o seu genoma oferece uma oportunidade única para identificar mudanças genéticas específicas a humanos completamente modernos do ponto de vista anatómico. Identificámos um fóssil com 38,000 anos de neandertal que estava excepcionalmente livre de contaminação de ADN de humanos modernos. A sequenciação directa de alta cadência de uma amostra de ADN deste fóssil forneceu até a este momento mais de um milhão de pares de bases de sequências de ADN nuclear hominóides. A comparação com os genomas humano e do chimpanzé revela que as sequências de ADN dos humanos modernos e dos neandertais divergiram em média 500,000 anos atrás. Os recursos fósseis e a tecnologia existente são neste momento suficientes para iniciar um esforço de sequenciação do genoma do neandertal.
O ênfase a negrito a meu e indica um facto muito importante a respeito desta amostra. O ADN recuperado do fóssil estava partido em inúmeros fragmentos com umas poucas centenas de pares de bases. Esses fragmentos foram tratados numa espécie de emulsão em que cada fragmento de ADN se assoc ia a uma espécie de conta oleosa suspensa em água. O procedimento de identificação do material na amostra, o chamado passo de pirossequenciação, é fantástico mas ficará para outra altura. As minhas últimas contribuições têm sido gigantescas e tenho perdido leitores. Digamos só que envolve fibras ópticas, luciferase (a enzima que faz os vagalumes brilharem) e um CCD para apanhar a luz.
Num só varrimento do método de sequenciação os cientistas são capazes de recuperar as sequências de centenas de milhar de fragmentos ao mesmo tempo. O que é uma boa coisa pois a maior parte do material genético é ADN de microorganismos que colonizaram o osso, ou que de alguma forma contaminaram a amostra. Esse ADN dos microorganismos é um problema, mas como temos o genoma completo do homem moderno poder ser identificado como muito diferente do humano e excluído da análise. Só que a parecença do ADN do neandertal com os humanos modernos coloca um problema. O fragmento de osso estudado foi manuseado por seres humanos modernos, logo está contaminado com ADN de Homo sapiens. O problema é sério. Em geral mesmo depois de os cientistas lavarem os fragmentos de osso, eliminarem a superfície, trabalharem em ambientes esterilizados e controlados, retirarem apenas amostras do anterior do osso, ainda assim encontram sempre ADN de humanos modernos, em quantidades impressionantes.
Para avaliarem a contaminação os cientistas deste estudo analisaram 70 amostras de ossos e dentes de neandertal. Desses 70 escolheram seis em melhores condições em que procuraram o ADN não do núcleo mas sim das mitocôndrias, usando amostras de 100 a 200 miligramas de osso. As sequências de ADN mitocondrial (ADNmt) dos neandertias são muito semelhantes mais ainda assim fáceis de distinguir do ADNmt do homem moderno. Os resultados foram os seguintes:Enquanto apenas cerca de 1% do ADNmt presente nas três amostras da França, Rússia e Uzbequistão eram do tipo-neandertal uma amostra da Croácia e uma de Espanha continham 5% e 75% de ADNmt tipo-neandertal. Um osso (Vi-80) da gruta de Vindija na Croácia, destacou-se por possuir ADMmt de neandertal em aproximadamente 99% dos segmentos com 63 pares de base e aproximadamente 94% dos segmentos de 119 pares de bases,
Claro que foi este Vi-80 a ser utilizado neste estudo, pois, admitindo que a frequência de contaminação é a mesma para o ADN nuclear, é uma amostra quase limpa.
Não vou discutir os resultados relativos a comparações com o genoma do chimpazé e dos humanos modernos, pois embora pareça muito um milhão de pares de bases é na verdade muito pouco. Faltam 3 ordens de grandeza mais para ter o genoma completo. Os autores estão no entanto optimistas e dão mesmo um prazo curto para chegar lá:Os resultados apresentados aqui foram obtidos a partir de aproximadamente um quinze avos de uma amostra preparada com cerca de 100 miligramas de osso. Para conseguir a cobertura completa do genoma neanderal (3 gigabases) sem melhorias na tecnologia, seriam necessários 20 gramas de osso e 6,000 varrimentos da versão corrente da plataforma 454 de sequenciação. Embora no presente essa seja uma tarefa avassaladora, podem facilmente antever-se melhorias técnicas nos procedimentos descritos aqui que fariam a recuperação de sequências de ADN cerca de 10 vezes mais eficiente (resultados nossos não publicados). Com essa perspectiva em vista, iniciámos recentemente um projecto que visa conseguir uma versão preliminar do genoma neandertal no prazo de dois anos.
Ou seja, é para breve.
Ficha técnica
Imagem do neandertal obtida no Neanderthaler Museum.
Referências
(ref1) Green, R. E. et al. (2006). Analysis of one million base pairs of Neanderthal DNA. Nature 444, 330–336. Laço DOI.
Ciência à distância de um clique
![]() O Adilson de Oliveira, responsável pelo blog por dentro da ciência, é um dos editores e colunistas duma nova experiência de divulgação científica, a revista digital clickCiência. O projecto conta com vários apoios mas está ligado essencialmente à Universidade Federal de São Carlos (Brasil). O lançamento formal pelos vistos só ocorrerá no dia 29 deste mês mas as páginas já estão a funcionar. Pelo que percebi é uma revista algo generalista, mas que em cada número coloca o destaque num tema em particular. No primeiro número o ênfase é nas nanotecnologias, embora faça também uma incursão astronómica, com uma coluna dedicada à despromoção de Plutão. As páginas são graficamente agradáveis, mas mantendo uma saudável sobriedade. Gosto particularmente do tucano que aparece no cabeçalho. Portanto já sabem, se querem saber mais sobre nanotecnologia, e para que serve, directamente do teclado dos especialistas, basta dar um saltinho ao clickCiência.
O Adilson de Oliveira, responsável pelo blog por dentro da ciência, é um dos editores e colunistas duma nova experiência de divulgação científica, a revista digital clickCiência. O projecto conta com vários apoios mas está ligado essencialmente à Universidade Federal de São Carlos (Brasil). O lançamento formal pelos vistos só ocorrerá no dia 29 deste mês mas as páginas já estão a funcionar. Pelo que percebi é uma revista algo generalista, mas que em cada número coloca o destaque num tema em particular. No primeiro número o ênfase é nas nanotecnologias, embora faça também uma incursão astronómica, com uma coluna dedicada à despromoção de Plutão. As páginas são graficamente agradáveis, mas mantendo uma saudável sobriedade. Gosto particularmente do tucano que aparece no cabeçalho. Portanto já sabem, se querem saber mais sobre nanotecnologia, e para que serve, directamente do teclado dos especialistas, basta dar um saltinho ao clickCiência.
sábado, novembro 25, 2006
A vida interior de uma célula, o musical
 Esta imagem de uma molécula a percorrer tranquilamente um filamento no interior de uma célula foi retirado de um filme que mostra o turbilhão de actividade que ocorre constantemente dentro das células. Tudo acompanhado por música. Vale a pena ver, é estranhamente relaxante. Encontrei esta referência no blog de uma amiga minha que está a passar um mau bocado. Vão até ao Living the Scientific Life para ver o filme.
Esta imagem de uma molécula a percorrer tranquilamente um filamento no interior de uma célula foi retirado de um filme que mostra o turbilhão de actividade que ocorre constantemente dentro das células. Tudo acompanhado por música. Vale a pena ver, é estranhamente relaxante. Encontrei esta referência no blog de uma amiga minha que está a passar um mau bocado. Vão até ao Living the Scientific Life para ver o filme.
Se passarem por lá vejam também uma outra contribuição com fantásticas imagens de fetos de mamíferos no útero.
Paquiceto, o mais antigo da linhagem

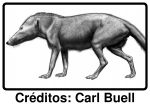 Esta é uma reconstrução de um Pakicetus, membro de um grupo de animais que viveram há mais de 50 milhões de anos na região que hoje é o Paquistão e Noroeste da Índia. Eram parentes próximos do antepassado de um grupo de animais de que falei aqui recentemente. Do tamanho de um lobo, possuíam pernas compridas, uma cauda robusta, e pequenos cascos a cobrirem as extremidades dos dedos, denunciando o seu parentesco com os artiodáctilos. Pelo aspecto, a afinidade com os seus parentes modernos mais próximos não é facil de estabelecer. Na verdade a relação entre o Pakicetus e as criaturas existentes hoje em dia repousa num conjunto de especializações do seu ouvido-interno. Quais são esses parentes modernos? Bem, o nome dá uma pista, pois indica não só o local de origem, Paki para Paquistão, mas também o grupo de mamíferos de que faz parte. [... ler mais]
Esta é uma reconstrução de um Pakicetus, membro de um grupo de animais que viveram há mais de 50 milhões de anos na região que hoje é o Paquistão e Noroeste da Índia. Eram parentes próximos do antepassado de um grupo de animais de que falei aqui recentemente. Do tamanho de um lobo, possuíam pernas compridas, uma cauda robusta, e pequenos cascos a cobrirem as extremidades dos dedos, denunciando o seu parentesco com os artiodáctilos. Pelo aspecto, a afinidade com os seus parentes modernos mais próximos não é facil de estabelecer. Na verdade a relação entre o Pakicetus e as criaturas existentes hoje em dia repousa num conjunto de especializações do seu ouvido-interno. Quais são esses parentes modernos? Bem, o nome dá uma pista, pois indica não só o local de origem, Paki para Paquistão, mas também o grupo de mamíferos de que faz parte. [... ler mais]
Pois é, o cetus é de cetáceo. Trata-se dos mais antigos cetáceos conhecidos. Pode parecer estranho que os cetáceos incluam animais com pernas e cascos, mas há 50 milhões de anos era assim. Este animal para todos os efeitos pode ser considerado uma baleia, Não vivia no oceano, e embora pudesse seguramente nadar, não seria um nadador extraordinário. Deambularia provavelmente pela foz ou estuários dos rios da região onde os seus restos foram encontrados.
A razão que me leva a falar neste animal não é nenhum artigo acabado de sair, mas mais uma excelente contribuição no blog de Carl Buell, que anunciou uma série de artigos e ilustrações sobre este tema. Para além de uma versão actualizada e a cores do Pakicetus há ilustrações de cetáceos modemos. Vão ao Olduvai George e fiquem maravilhados. O mais extraordinário é a promessa de que há mais a caminho.
Ficha técnica
Ilustração de Pakicetus cortesia de Carl Buell, retirada desta página. Esta imagem pode ser utilizada livremente desde que inclua a legenda: Illustration by Carl Buell, and taken from http://www.neoucom.edu/Depts/Anat/Pakicetid.html, ou uma tradução em português que foi o que fiz.
Assobiar como um tucuxi ou latir como um boto




 Por vezes as águas de Mamirauá são cortadas por uma grande e robusta barbatana dorsal, que pertence a um golfinho conhecido localmente como tucuxi, de seu nome científico Sotalia fluviatilis. Para perceber a graciosidade destas criaturas, nada melhor que vê-las em filme, cortesia do ARKive. Como os restantes golfinhos, o tucuxi recorre à capacidade de ecolocalização, usando estalidos de alta de alta frequência (por volta dos 150 kHz), para se orientar e navegar. Mas as capacidades vocais dos tucuxis, e dos golfinhos em geral, não se esgotam na ecolocalização, emitem também vocalizações a frequências bastante mais baixas, vocalizações essas que têm um papel importante na sua vivência ecológica e social. Uma das vocalizações que tem sido motivo de grande interesse são os assobios tonais. Uma das questões acerca destes assobios é a sua origem evolutiva. Serão uma inovação, que apareceu apenas num grupo de golfinhos oceânicos, os chamados delfinídeos, ou serão uma modificação de um mecanismo ancestral? [... ler mais]
Por vezes as águas de Mamirauá são cortadas por uma grande e robusta barbatana dorsal, que pertence a um golfinho conhecido localmente como tucuxi, de seu nome científico Sotalia fluviatilis. Para perceber a graciosidade destas criaturas, nada melhor que vê-las em filme, cortesia do ARKive. Como os restantes golfinhos, o tucuxi recorre à capacidade de ecolocalização, usando estalidos de alta de alta frequência (por volta dos 150 kHz), para se orientar e navegar. Mas as capacidades vocais dos tucuxis, e dos golfinhos em geral, não se esgotam na ecolocalização, emitem também vocalizações a frequências bastante mais baixas, vocalizações essas que têm um papel importante na sua vivência ecológica e social. Uma das vocalizações que tem sido motivo de grande interesse são os assobios tonais. Uma das questões acerca destes assobios é a sua origem evolutiva. Serão uma inovação, que apareceu apenas num grupo de golfinhos oceânicos, os chamados delfinídeos, ou serão uma modificação de um mecanismo ancestral? [... ler mais]
Para perceber os termos que vou utilizar em seguida, sugiro, aos que não o fizeram ainda, a leitura da minha contribuição sobre a filogenia dos golfinhos, pois convém ter presente o cladograma que aí apresento. O tucuxi pertence a um dos grupos desse cladograma, os delfinídeos, que se caracterizam pelos tais assobios tonais; apenas 4 das 32 espécies do grupo não os utilizam. Ser tonal significa concentrar a maior parte da energia acústica numa faixa de frequências muito estreita. Eis em baixo um exemplo da intensidade das vocalizações, em função da frequência da emissão, para o tucuxi: Notem bem a emissão concentrada numa estreita banda que sobe gradualmente em frequência até estabilizar numa frequência ligeiramente abaixo dos 20 kHz. Este é o tipo de representação das propriedades acústicas que define um assobio.
Notem bem a emissão concentrada numa estreita banda que sobe gradualmente em frequência até estabilizar numa frequência ligeiramente abaixo dos 20 kHz. Este é o tipo de representação das propriedades acústicas que define um assobio.
Outros golfinhos, como por exemplo as belugas, conhecidas como "os canários do oceano", utilizam abundamente os seus dotes vocais, mas não recorrem a nada com um carácter tonal tão marcado. Na verdade, nenhum dos membros dos Phocoenidae ou dos Monondontidae parece incluir os "assobios" no seu reportório vocal. Isso levou alguns autores a sugerirem que os assobios seriam uma inovação associada apenas aos delfinídeos. Ora os botos-vermelhos (Inia geoffrensis) também produzem vocalizações. Se as vocalizações dos botos-vermelhos fossem homólogas às dos delfinídeos, dadas as relações de parentesco mais distantes dos botos-vermelhos com os restantes grupos de golfinhos, poder-se-ia avançar com uma hipótese algo diferente: os assobios seriam algo que existiria num antepassado comum aos botos e restantes golfinhos, e que teria sido perdida posteriormente durante a evolução das belugas, narvais e focenídeos.
Para clarificar o tipo de vocalização efectivamente produzida pelos botos, Jeffrey Podos, Vera da Silva e Marcos Rossi-Santos discutem num estudo publicado na revista Ethology (ref1) os resultados de cerca de 40 horas de registos sonoros obtidos no lago Mamirauá. Numa tradução livre do resumo:Os golfinhos oceânicos (Odontoceti: Delphinidae) produzem assobios tonais, cuja estrutura e função são razoavelmente bem caracterizadas. Bastante menos se conhece sobre as origens evolutivas dos assobios dos golfinhos, incluindo informação básica sobre a estrutura vocal em grupos taxonómicos irmãos como os golfinhos fluviais dos Platanistidae. Caracterizamos aqui as vocalizações do golfinho do Rio Amazonas (Inia geoffrensis), para o qual foram relatados assobios mas que não foram bem documentados. Estudámos Inia na Reserva de Desenvolvimento Sustentável Mamirauá na Amazónia Brasileira. Durante 480 blocos de 5 minutos (mais de 5 semanas) monitorizámos e gravámos vocalizações, anotámos o tamanho do grupo e a actividade, e registámos as subidas à superfície para respiração e para mergulho. Globalmente, a produção vocal dos Inia estava correlacionada positivamente com as subidas para mergulho, sugerindo que as vocalizações estão associadas com a alimentação. As análises acústicas mostraram que as vocalizações do Inia são estruturalmente distintas dos assobios delfinídios simples, incluindo aqueles do delfinídio Sotalia fluviatilis registado no nosso local. Estes dados apoiam a hipótese de que os assobios são uma vocalização derivada única aos delfinídeos.
O resumo é algo curto, e alguns dos pontos merecem ser apresentados com um pouco mais de detalhe. Jeffrey Podos e colegas referem que os botos se mostraram muito comedidos, diga-se mesmo avaros, nas suas vocalizações. Cada boto-vermelho só vocalizava em média uma vez a cada 10.1 minutos, e em 40% dos blocos de 5 minutos não foram registadas quaisquer vocalizações. Os autores referem mesmo que cerca de metade das vocalizações dos botos foram produzidas em apenas 10% dos blocos. Contrastando com esta escassez sonora, os tucuxis foram observados a assobiar em 94% das vezes em que foram detectados.
Quanto ao que os botos estavam a fazer nos períodos em que vocalizavam, os autores do artigo, que não podiam observar abaixo da água, pois era bastante turva, conseguiram algumas pistas pela forma como os botos ascendiam à superfície. Distinguiram duas formas distintas. Uma era a subida horizontal ou para respiração. Nesta tipo de subidas o animal trazia a cabeça à tona, ou ascendia com a cabeça e tronco alinhados: O outro tipo de idas às superfície eram as subidas enroladas ou de mergulho. Durante as subidas enroladas, os golfinhos normalmente expunham apenas o dorso, contraíam o corpo numa forma em 'c' e rapidamente deslizavam para a frente. No artigo não vinham ilustrações mas presumo que seja mais ou menos isto:
O outro tipo de idas às superfície eram as subidas enroladas ou de mergulho. Durante as subidas enroladas, os golfinhos normalmente expunham apenas o dorso, contraíam o corpo numa forma em 'c' e rapidamente deslizavam para a frente. No artigo não vinham ilustrações mas presumo que seja mais ou menos isto: Os autores verificaram que as vocalizações dos botos-vermelhos estavam correlacionadas com as subidas enroladas e não com as horizontais. Os botos vermelhos executam as subidas enroladas quando empreendem mergulhos profundos, o que normalmente ocorre quando se alimentam. O comportamento vocal dos botos está assim associado temporalmente com a alimentação.
Os autores verificaram que as vocalizações dos botos-vermelhos estavam correlacionadas com as subidas enroladas e não com as horizontais. Os botos vermelhos executam as subidas enroladas quando empreendem mergulhos profundos, o que normalmente ocorre quando se alimentam. O comportamento vocal dos botos está assim associado temporalmente com a alimentação.
Quando se analisa a intensidade das vocalizações como função de frequência a que são emitidas, vê-se claramente que as emissões do boto são muito diferentes das do tucuxi. Eis um exemplo de emissões de um boto: Além de muito mais diversificadas, consistindo em séries de notas de muito curta duração, ocorrem a frequências muito mais baixas, e mostram estrutura harmónica. Por estrutura harmónica entende-se a existência de bandas como o mesmo comportamento temporal mas a frequências múltiplas, ou seja as "emissões semelhantes umas em cima das outras" que se veêm nos gráficos acima.
Além de muito mais diversificadas, consistindo em séries de notas de muito curta duração, ocorrem a frequências muito mais baixas, e mostram estrutura harmónica. Por estrutura harmónica entende-se a existência de bandas como o mesmo comportamento temporal mas a frequências múltiplas, ou seja as "emissões semelhantes umas em cima das outras" que se veêm nos gráficos acima.
Os autores falam um pouco mais sobre as diferenças e possíveis utilizações das vocalizações do boto e dos delfinídeos no final do artigo:As funções dos assobios dos golfinhos começaram a ser elucidades. Por exemplo, surgem na comunicação entre mãe e cria, e interações nos grupos. O nosso estudo mostro-se pouco esclarecedor sobre a função do comportamento vocal dos Inia. Poderão ser utilizadas para facilitar a pesca cooperativa ou serem apenas um efeito colateral sem nenhuma função particular. Não se encontraram indícios de outros tipos de utilização. Por exemplo, mãe e crias não vocalizam quando se reunem após uma separação temporária, e não foram registadas respostas ao passar de vocalizações gravadas.
Ou seja, há ainda muito que estudar.
Há ainda outro aspecto a merecer a atenção dos autores: poderão as vocalizações dos botos ser um precursor dos assobios dos delfinídeos? Jeffrey Podos e colegas pensam que não. Um dos argumentos que avançam é que o roaz corvineiro (Tursiops truncatus), um delfinídeo, produz vocalizações, chamadas "latidos", que são muito semelhantes às dos botos. O mecanismo que as produz deverá por isso ter sido conservado do ponto de vista evolutivo. Mas a diferença mais importante tem a ver com a forma como a frequência no máximo das emissões varia com o tamanho dos animais. Isso é algo que se mostra na figura abaixo para o boto-vermelho (triângulo) e várias espécies de delfinídios (círculos). É óbvio o grande afastamento do boto em relação à linha que reproduz o comportamento médio dos delfinídeos. Segundo Jeffrey Podos e colegas este desvio é uma idicação muito forte de que as vocalizações no boto são produzidas por um mecanismo distinto, embora coloquem uma ressalva: conhece-se muito pouco sobre os mecanismos de produção de som nos golfinhos. A fonte dos assobios permanece por descobrir. Mostrar que os "latidos" do boto e os "assobios" do tucuxi provêm de fontes sonoras distintas seria a forma mais adequada de mostrar que não são homólogos.
É óbvio o grande afastamento do boto em relação à linha que reproduz o comportamento médio dos delfinídeos. Segundo Jeffrey Podos e colegas este desvio é uma idicação muito forte de que as vocalizações no boto são produzidas por um mecanismo distinto, embora coloquem uma ressalva: conhece-se muito pouco sobre os mecanismos de produção de som nos golfinhos. A fonte dos assobios permanece por descobrir. Mostrar que os "latidos" do boto e os "assobios" do tucuxi provêm de fontes sonoras distintas seria a forma mais adequada de mostrar que não são homólogos.
Para já despeço-me do boto-vermelho, mas ficarei atento a novidades. Voltarei no entanto em breve aos delfinídeos, para falar nas funções dos assobios. Se, como tudo parece apontar, se trata de uma característica única dos delfinídeos, para que serve e o que despoletou essa inovação?
Ficha técnica
Imagens "fotográficas" retirada das páginas do Projeto Boto.
Restantes figuras e inspiração para o texto retiradas do artigo abaixo (ref1).
Referências
(ref1) Jeffrey Podos, Vera M. F. da Silva & Marcos R. Rossi-Santos (2002). Vocalizations of Amazon River Dolphins, Inia geoffrensis: Insights into the Evolutionary Origins of Delphinid Whistles. Ethology 108, 601-612. Laço DOI.
sexta-feira, novembro 24, 2006
A importância de ter pernas curtas quando lagarto come lagarto

 Esta contribuição não tem a ver com os eventos futebolísticos de quarta-feira. Aliás os lagartos de que vou falar nem sequer equipam de verde. A criatura da imagem é um anole castanho, de seu nome científico Anolis sagrei. Trata-se de um animal endémico das Baamas que coloniza algumas pequenas ilhotas da região. Os anoles são bons trepadores e podem subir a ramos e superfícies lisas, mas encontram-se também muitas vezes no chão. São notáveis corredores e numa ilha livre de predadores os animais com maiores capacidades atléticas (pernas maiores) têm alguma vantagem sobre os animais mais lentos. Ora alguns cientistas resolveram colocar nalgumas dessas ilhas um outro lagarto, a iguanita de rabo enroscado, Leiocephalus carinatus, e ver o que se passava. É que a iguanita também é boa corredora e não desdenha engolir qualquer anole que se meta a jeito. [... ler mais]
Esta contribuição não tem a ver com os eventos futebolísticos de quarta-feira. Aliás os lagartos de que vou falar nem sequer equipam de verde. A criatura da imagem é um anole castanho, de seu nome científico Anolis sagrei. Trata-se de um animal endémico das Baamas que coloniza algumas pequenas ilhotas da região. Os anoles são bons trepadores e podem subir a ramos e superfícies lisas, mas encontram-se também muitas vezes no chão. São notáveis corredores e numa ilha livre de predadores os animais com maiores capacidades atléticas (pernas maiores) têm alguma vantagem sobre os animais mais lentos. Ora alguns cientistas resolveram colocar nalgumas dessas ilhas um outro lagarto, a iguanita de rabo enroscado, Leiocephalus carinatus, e ver o que se passava. É que a iguanita também é boa corredora e não desdenha engolir qualquer anole que se meta a jeito. [... ler mais]
Os resultados desde estudo em selecção "natural" foram publicados num artigo muito curto de Jonathan Losos e colegas na revista Science (ref1). Numa tradução livre do resumo:Serão as espécies capazes de se adaptar quando o ambiente muda? Ao conduzir experiências em ambientes naturais, os biólogos podem estudar a forma como processos evolucionários tais como a selecção natural operam através do tempo. Nós previmos que a introdução de um predador terrestre iria primeiro seleccionar lagartos de pernas mais compridas, que são mais rápidos, mas à medida que os lagartos se deslocavam para ramagens mais elevadas para evitar o predador, a selecção iria inverter-se e favorecer os indivíduos de pernas curtas que se movimentam melhor aí. A nossa experiência em 12 ilhotas confirmou estas previsões em apenas uma geração, mostrando assim a velocidade com que as forças evolucionárias podem mudar durante tempos de fluxo ambiental.
O que os autores fizeram foi estudar 12 ilhas, 6 em que introduziram predadores, e 6 em que não o fizeram, para servir de controlo. Nessas ilhas os autores verificaram o tamanho das patas e o tempo que os animais passavam nos ramos antes da introduzirem as iguanitas, 6 meses depois de o fazerem e um ano depois de os predadores terem feito a sua aparição. Seis meses após o início da experiência, a média do tamanho das patas dos sobreviventes era maior, ou seja mais pernas curtas tinham sido comidos, mas 12 meses depois a média do tamanho das patas era substancialmente menor, os pernas longas tinham sido comidos. Os pernas curtas sobreviventes tinham maior facilidade em trepar às plantas que os penas longas e evitavam mais facilmente os lagartos com o rabo enrolado.
O tal estádio intermédio de que falam os autores é um pouco como aquela velha anedota: dados dois indivíduos a fugirem de um urso, um que queira escapar não tem que se preocupar em correr mais que o urso, basta-lhe correr mais que o outro. As iguanitas ao correrem atrás dos anoles no chão comeram primeiro os lentos e, quando eles se tornaram raros, os rápidos. Deve no entanto notar-se que a chave para a sobrevivência a longo termo foi outra: os autores do artigo referem que houve uma mudança de comportamento dos anoles sobreviventes. Não escaparam apenas os que por algum tipo de predisposição inata passavam já mais tempo nos ramos antes da invasão. Considerando apenas os anoles que escaparam, antes da invasão esses animais passavam 60% do tempo empoleirados em ramos, após a invasão passavam 90%. Os anoles que escaparam não eram nada burros.
Esta é sem dúvida uma experiência muito interessante em aptidão evolutiva, e como ela pode mudar rapidamente. Se deixássemos estes ecossistemas evoluírem durante milénios, sem mais interferências, é provável que a tendência se acentuasse. Ou talvez não, é que estas ilhotas são varridas periodicamente por furacões. Esse aspecto está ligado a algo que, desde o início, me pareceu curioso nesta história: o aspecto ético da coisa. Aposto que a maior parte dos leitores não se sentiu muito incomodada pelo destino dos anoles, afinal são "apenas" lagartos. No entanto muitos considerariam uma crueldade se se tratasse de uma população de aves. Após pesquisar um bocado descobri que na verdade o que os autores fizeram foi apenas "apressar" um pouco a natureza. As ilhotas têm qualquer coisa como uns meros 750 metros quadrados cada, estão praticamente ao nível do mar, e ficam a pouco mais de 100 metros das massas de terra maior. As iguanitas devoradoras de anoles invadem-nas com frequência. As pequenas ilhas são no entanto varridas frequentemente por furacões, e as populações de iguanitas residentes, e muitas vezes mesmo as de anoles, têm uma tendência para se afogarem, e se extinguirem até à invasão seguinte. Daí que essas ilhotas estejam frequentemente livres de predadores.
Os anoles parecem-se vagamente com lagartixas mas os machos possuem uma bela papada em leque, colorida com uns garridos tons de laranja e amarelo, que usam como uma espécie de sinal para marcar o território. Merecem mais uma foto. Ficha técnica
Ficha técnica
Imagem do anole no ínicio da contribuição cortesia de Alison Phillips, retirada desta página da Wikimedia Commons.
Outra imagem de anole cortesia de Stephen Lea, retirada desta página da Wikimedia Commons.
Referências
(ref1) Jonathan B. Losos, Thomas W. Schoener, R. Brian Langerhans, David A. Spiller (2006). Rapid Temporal Reversal in Predator-Driven Natural Selection. Science, Vol. 314. no. 5802, p. 1111. Laço DOI.
quinta-feira, novembro 23, 2006
Unicórnios, baleias brancas e assassinas, a espantosa diversidade dos golfinhos

 Estas imagens mostram narvais, de seu nome científico Monodon monoceros. Embora a afiada coisa que se projecta da cabeça não se trate exactamente de um corno, mas sim de um dente, este animal não deixa de evocar o mitológico unicórnio. É uma criatura com o seu quê de mágico. O narval é um bom exemplo da diversidade dos seres incluídos sob a designação de golfinhos. Quando se analisa essa variedade conseguimos contudo distinguir um certo número de grupos naturais dentro dos golfinhos: alguns golfinhos são mais iguais a certos golfinhos que a outros. Esses grupos naturais partilham entre si características de dentição, morfologia, e comportamento, entre outras. Isso é um indicador de que as espécies no grupo possuem um antepassado comum entre elas que não partilham com os membros dos outros grupos. As relações de parentesco são importantes para compreender a evolução de muitas características da ecologia e comportamento destes animais. Resolvi por isso, antes de introduzir o tema das vocalizações, fazer um pequeno parêntesis para abordar as relações familiares entre as muitas espécies de golfinhos e seus "aliados". [... ler mais]
Estas imagens mostram narvais, de seu nome científico Monodon monoceros. Embora a afiada coisa que se projecta da cabeça não se trate exactamente de um corno, mas sim de um dente, este animal não deixa de evocar o mitológico unicórnio. É uma criatura com o seu quê de mágico. O narval é um bom exemplo da diversidade dos seres incluídos sob a designação de golfinhos. Quando se analisa essa variedade conseguimos contudo distinguir um certo número de grupos naturais dentro dos golfinhos: alguns golfinhos são mais iguais a certos golfinhos que a outros. Esses grupos naturais partilham entre si características de dentição, morfologia, e comportamento, entre outras. Isso é um indicador de que as espécies no grupo possuem um antepassado comum entre elas que não partilham com os membros dos outros grupos. As relações de parentesco são importantes para compreender a evolução de muitas características da ecologia e comportamento destes animais. Resolvi por isso, antes de introduzir o tema das vocalizações, fazer um pequeno parêntesis para abordar as relações familiares entre as muitas espécies de golfinhos e seus "aliados". [... ler mais]
Comecemos então com um velho conhecido nosso. Os botos-vermelhos, Inia geoffrensis e as toninhas de rio sul-americanas, Pontoporia blainvillei, formam um grupo à parte do resto. Existem muitas características neste grupo que os restantes golfinhos não têm, e vice-versa. O "resto" dos golfinhos pode separar-se por sua vez em três ramos bem distintos. Um desses ramos são as três espécies dos chamados Monodontidae. Os monodontídeos tiram o seu nome do narval (Monodon monoceros) e incluem ainda as belugas (Delphinapterus leucas). Já mostrei aqui uma imagem dos fabulosos unicórnios marinhos no início da contribuição. As simpáticas baleias-branca, que na verdade são golfinhos, também merecem uma imagem:
Os botos-vermelhos, Inia geoffrensis e as toninhas de rio sul-americanas, Pontoporia blainvillei, formam um grupo à parte do resto. Existem muitas características neste grupo que os restantes golfinhos não têm, e vice-versa. O "resto" dos golfinhos pode separar-se por sua vez em três ramos bem distintos. Um desses ramos são as três espécies dos chamados Monodontidae. Os monodontídeos tiram o seu nome do narval (Monodon monoceros) e incluem ainda as belugas (Delphinapterus leucas). Já mostrei aqui uma imagem dos fabulosos unicórnios marinhos no início da contribuição. As simpáticas baleias-branca, que na verdade são golfinhos, também merecem uma imagem: Um outro grupo natural de golfinhos os Phocoenidae, inclui uma espécie razoavelmente comum no litoral português, que pode ser visto por exemplo em Tróia e na Arrábida. De seu nome científico Phocoena phocoena, é conhecido localmente por boto, ou seja o mesmo nome dado ao seu parente afastado do Amazonas.
Um outro grupo natural de golfinhos os Phocoenidae, inclui uma espécie razoavelmente comum no litoral português, que pode ser visto por exemplo em Tróia e na Arrábida. De seu nome científico Phocoena phocoena, é conhecido localmente por boto, ou seja o mesmo nome dado ao seu parente afastado do Amazonas. Aliás, para mostrar quão enganador pode ser um nome, este animal é também conhecido em Portugal por toninha, que é a designação dada no Brasil ao Pontoporia blainvillei. Os focenídeos são também um grupo pequeno, incluem apenas seis espécies.
Aliás, para mostrar quão enganador pode ser um nome, este animal é também conhecido em Portugal por toninha, que é a designação dada no Brasil ao Pontoporia blainvillei. Os focenídeos são também um grupo pequeno, incluem apenas seis espécies.
O grupo maior, com 32 espécies, e que inclui os golfinhos mais comuns, são os Delphinidae. Os delfinídeos incluem o mais famoso golfinho de todos, o roaz-corvineiro, Tursiops truncatus. Trata-se um golfinho que se adapta bem ao cativeiro e ao convívio com humanos. Eis aqui uma imagem de um roaz-corvineiro de nome K-Dog numa missão de desminagem no Golfo Pérsico. Para aqueles que estão a pensar comentar o facto de eu estar a fazer propaganda de uma incursão militar altamente controversa, devo dizer que a missão deste golfinho tinha cariz não bélico: destinava-se a manter desimpedida uma rota usada por barcos de ajuda humanitária. Hesitei um bocado, mas é uma fotografia excelente, e de divulgação livre.
Para aqueles que estão a pensar comentar o facto de eu estar a fazer propaganda de uma incursão militar altamente controversa, devo dizer que a missão deste golfinho tinha cariz não bélico: destinava-se a manter desimpedida uma rota usada por barcos de ajuda humanitária. Hesitei um bocado, mas é uma fotografia excelente, e de divulgação livre.
Um outro delfinídeo bastante conhecido é a infame baleia-assassina, Orcinus orca que, mais uma vez, apesar do nome baleia, é um golfinho. Agora que já mostrei os vários grupos e apresentei imagens, vou então falar brevemente das relações de parentesco. A árvore filogenética correspondente, construída com base nas características das várias espécies de golfinhos, é a seguinte:
Agora que já mostrei os vários grupos e apresentei imagens, vou então falar brevemente das relações de parentesco. A árvore filogenética correspondente, construída com base nas características das várias espécies de golfinhos, é a seguinte: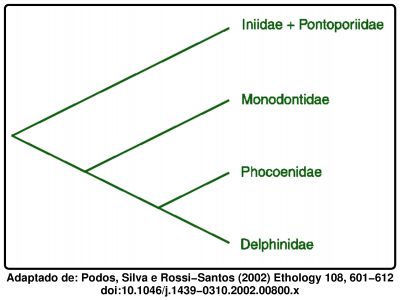 Nesta representação, designada por cladograma (do grego clados para ramo), cada união entre ramos significa que os grupos em questão partilham um antepassado comum que não partilham com os grupos mais abaixo. Este cladograma indica que os Delphinidae, Phoecoenidae e Monodontidae partilham entre si um antepassado comum mais recente do que qualquer um deles partilha com os boto-vermelhos e toninhas sul-americanos (Iniidae e Pontoporiidae). Por sua vez os delfinídeos e focenídeos partilham entre si um antepassado comum mais recente do que qualquer um deles partilha com as belugas e narvais.
Nesta representação, designada por cladograma (do grego clados para ramo), cada união entre ramos significa que os grupos em questão partilham um antepassado comum que não partilham com os grupos mais abaixo. Este cladograma indica que os Delphinidae, Phoecoenidae e Monodontidae partilham entre si um antepassado comum mais recente do que qualquer um deles partilha com os boto-vermelhos e toninhas sul-americanos (Iniidae e Pontoporiidae). Por sua vez os delfinídeos e focenídeos partilham entre si um antepassado comum mais recente do que qualquer um deles partilha com as belugas e narvais.
Esta discussão da filogenia foi apenas uma introdução simplificada, destinada à contribuição sobre as vocalizações dos golfinhos que apresentarei em seguida. Voltarei eventualmente a este tema, de forma mais elaborada, discutindo toda a árvore filogenética dos cetáceos, que, por incrível que pareça, são aparentados aos artiodáctilos, o grupo que inclui, entre outros animais, hipopótamos, camelos e porcos.
Ficha técnica
Imagens de narvais cortesia de Kristin Laidre, retiradas desta página do OceanExplorer da NOAA.
Imagem da beluga cortesia de Robyn Angliss, retirada desta página do National Marine Mammal Laboratory.
Imagem da orca e da Phocoena phocoena retiradas a partir desta página da Marine Mammal Commission.
Imagem do boto-vermelho retirada das páginas do Projeto Boto.
Imagem do roaz-corvineiro cortesia de Brien Aho, retirada do US Navy newsstand.
segunda-feira, novembro 20, 2006
Quando para amar é preciso saber morder

 Eu tinha indicado na contribuição precedente que iria falar das vocalizações dos botos-vermelhos, Inia geoffrensis, mas resolvi deixar isso para depois, pois encontrei algo igualmente interessante. Esta imagem mostra um comportamento característico dos botos machos adultos durante a época do acasalamento: o uso de objectos durante paradas com carácter social e sexual. Mas esta é apenas a parte mais fácil do processo de acasalamento, os pobres machos têm que penar muito mais para terem sucesso na sua vida amorosa. Os botos não ficam passivos a verem outros botos desfilarem em frente aos alvos da sua afeição: aparentemente os machos agridem-se ferozmente uns aos outros, e as marcas disso são visíveis nos seus corpos. Este é mais um dos resultados só possíveis pela estudo de uma grande população destes animais que tem vido a ser feito há mais de dez anos na Reserva de Desenvolvimento Sustentável Mamirauá no âmbito do Projeto Boto. Os resultados são fascinantes e mostram que o boto é de facto um caso à parte nos cetáceos. [... ler mais]
Eu tinha indicado na contribuição precedente que iria falar das vocalizações dos botos-vermelhos, Inia geoffrensis, mas resolvi deixar isso para depois, pois encontrei algo igualmente interessante. Esta imagem mostra um comportamento característico dos botos machos adultos durante a época do acasalamento: o uso de objectos durante paradas com carácter social e sexual. Mas esta é apenas a parte mais fácil do processo de acasalamento, os pobres machos têm que penar muito mais para terem sucesso na sua vida amorosa. Os botos não ficam passivos a verem outros botos desfilarem em frente aos alvos da sua afeição: aparentemente os machos agridem-se ferozmente uns aos outros, e as marcas disso são visíveis nos seus corpos. Este é mais um dos resultados só possíveis pela estudo de uma grande população destes animais que tem vido a ser feito há mais de dez anos na Reserva de Desenvolvimento Sustentável Mamirauá no âmbito do Projeto Boto. Os resultados são fascinantes e mostram que o boto é de facto um caso à parte nos cetáceos. [... ler mais]
Este aspecto da vida dos botos é descrito em detalhe num estudo de Tony Martin e Vera da Silva na revista Marine Mammal Science (ref1). Numa tradução livre do resumo:Obtiveram-se medições e descrições quantitativas de uma grande amostra de botos (Inia geoffrensis) adultos vivos da Reserva de Mamirauá no Amazonas central. Os machos eram em média 16% maiores e pesavam 55% mais que as fêmeas, mostrando que esta espécie é uma das mais sexualmente dimórficas em tamanho de todos os cetáceos.
Esta diferença de tamanho não existe noutros golfinhos de rio, e entre os cetáceos verifica-se apenas nuns poucos como o cachalote (Physeter catodon), a beluga (Delphinapterus leucas) o narval (Monodon monoceros) e alguns delfinídeos como a orca (Orcinus orca). O resumo aborda em seguida a questão da cor e das cicatrizes:Os machos eram também mais rosados que as fêmeas, e mais fortemente cobertos de cicatrizes provocadas por filas de dentes da mesma espécie, e possuiam mais ferimentos com risco de vida.
Segundo referem os autores, no corpo do artigo, a cor rosada ocorre por um processo natural de envelhecimento da pele, em que o tom cinza passa a tons cada vez mais rosados, mas também tem a ver com as cicatrizes, que são rosa claro. Os autores especulam que o grau de coloração rosa pode ser usado como uma espécie de propaganda de maturidade por parte dos machos. Pois é, segundo parece nos botos "macho que é macho" veste cor de rosa.
Mas nos botos "macho que é macho" não se limita a vestir de cor de rosa, está também coberto de cicatrizes e sinais de violência. Eis aqui em baixo uma foto da cabeça de um boto macho adulto. Notem o padrão de pigmentação que mostra evidências de abrasão, como se o animal tivesse sido esfregado com lixa. Notem ainda o grande número de cicatrizes no animal. As barbatanas caudais de muitos animais mostram também uma história de confrontos. Todas estas estrias são marcas de dentes.
As barbatanas caudais de muitos animais mostram também uma história de confrontos. Todas estas estrias são marcas de dentes. Este tipo de "decoração" é quase universal nos botos, com a excepção de alguns animais muito jovens. A distância entre os dentes nas estrias mostra que não foram feitas por animais de outras espécies existentes na área. Os possíveis candidatos eram um outro golfinho, o tucuxi (Sotalia fluviatilis) e os jacarés (Melanosuchus niger e Caiman crocodilus). Mas não foram animais dessas espécies, estas marcas foram necessariamente infligidas por outros botos.
Este tipo de "decoração" é quase universal nos botos, com a excepção de alguns animais muito jovens. A distância entre os dentes nas estrias mostra que não foram feitas por animais de outras espécies existentes na área. Os possíveis candidatos eram um outro golfinho, o tucuxi (Sotalia fluviatilis) e os jacarés (Melanosuchus niger e Caiman crocodilus). Mas não foram animais dessas espécies, estas marcas foram necessariamente infligidas por outros botos.
As marcas que os botos deixam uns nos outros não se limitam a uns quantos arranhões. Nalguns casos os botos mostram mesmo danos estruturais nalgumas partes do seu corpo. Notem bem os pedaços que faltam nesta barbatana caudal. Os machos apresentam mesmo uma característica que pode ser uma adaptação destinada a esta vida de combate:
Os machos apresentam mesmo uma característica que pode ser uma adaptação destinada a esta vida de combate:Alguns machos maiores tinham áreas de pele modificada que podiam ser simplesmente tecido de cicatrização, mas que poderiam ser também uma característica hereditária usada como escudo ou arma.
Estas zonas encontram-se por exemplo nas barbatanas peitorais: e na região anterior à barbatana caudal:
e na região anterior à barbatana caudal: No artigo em inglês os autores referem-se a este tipo de lesões como cobblestone skin, que poderíamos traduzir à letra como pele de macadame. Apenas os machos as possuem. As fêmeas, mesmo quando apresentam cicatrizes nestas áreas, nunca mostram algo semelhante. Os autores avançam a hipótese que se trata de uma adaptação: uma espécie de calosidade que funcionará ou como escudo, ou com intuitos ofensivos para atacar oponentes.
No artigo em inglês os autores referem-se a este tipo de lesões como cobblestone skin, que poderíamos traduzir à letra como pele de macadame. Apenas os machos as possuem. As fêmeas, mesmo quando apresentam cicatrizes nestas áreas, nunca mostram algo semelhante. Os autores avançam a hipótese que se trata de uma adaptação: uma espécie de calosidade que funcionará ou como escudo, ou com intuitos ofensivos para atacar oponentes.
As cicatrizes são muito mais comuns entre machos e os danos realmente sérios também. Segundo os autores do artigo tem tudo a ver com a busca do amor:Tal como nos cachalotes, o dimorfismo sexual e a agressão entre machos parecem estar ligados nos botos, sugerindo uma competição feroz por um recurso -- provavelmente oportunidades de acasalamento. O boto é único entre os golfinhos de rio no facto de os machos serem maiores que as fêmeas. Esta distinção implica uma longa separação evolutiva e diferenças fundamentais de comportamento social.
Todos estes comportamentos são apenas inferidos. É preciso muito mais trabalho de campo para comprovar estas suposições. Na próxima contribuição irei encerrar esta "investida" nos domínios do boto-vermelho do Amazonas com aspectos relativos às vocalizações deste intrigante animal.
Ficha técnica
Imagem no início da contribuição retirada das páginas do Projeto Boto.
Restantes imagens e inspiração para o texto retiradas do artigo abaixo (ref1).
Referências
(ref1) Martin, A.R., da Silva, V.M.F. (2006). SEXUAL DIMORPHISM AND BODY SCARRING IN THE BOTO (AMAZON RIVER DOLPHIN) INIA GEOFFRENSIS. Marine Mammal Science 22(1): 17-24. Laço DOI.
sábado, novembro 18, 2006
Urubu d'água, o apetite colombiano que está a dizimar o boto

 O peixe da imagem é a piracatinga, Calophysus macropterus, um bicho essencialmente necrófago, que come os cadáveres de animais aquáticos, conhecido por isso pela designação urubu d'água. Com tal designação, e com tais hábitos alimentares, não espanta que não seja muito popular como alimento no Brasil, e até há pouco tempo a sua pesca não tinha grande expressão. É no entanto muito apreciado na Colômbia, onde é fatiado, transformado em filetes e vendido em Bogotá, ou então exportado para os Estados Unidos da América, onde o catfish é razoavelmente popular. Até aqui tudo bem, não há de errado em gostar de um alimento, por muito repugnante que pareça a outras pessoas. Além disso o urubu d'água é bastante comum em muitas partes da bacia do Amazonas, incluindo regiões do Brasil, por exemplo na área coberta pela Reserva de Desenvolvimento Sustentável Mamirauá. O apetite dos colombianos por esta criatura tornaram-na um bem relativamente escasso nesse país e a pesca atravessou fronteiras, para o lado brasileiro, essencialmente dedicada à exportação para a Colômbia. O efeito nos botos vermelhos tem sido devastador. Não se trata de problemas de golfinhos presos em redes, ou dos pescadores a tentarem livrar-se de um competidor indesejado. Não, é uma história de desperdício criminoso, revoltante, e mesmo com requintes de malvadez. [... ler mais]
O peixe da imagem é a piracatinga, Calophysus macropterus, um bicho essencialmente necrófago, que come os cadáveres de animais aquáticos, conhecido por isso pela designação urubu d'água. Com tal designação, e com tais hábitos alimentares, não espanta que não seja muito popular como alimento no Brasil, e até há pouco tempo a sua pesca não tinha grande expressão. É no entanto muito apreciado na Colômbia, onde é fatiado, transformado em filetes e vendido em Bogotá, ou então exportado para os Estados Unidos da América, onde o catfish é razoavelmente popular. Até aqui tudo bem, não há de errado em gostar de um alimento, por muito repugnante que pareça a outras pessoas. Além disso o urubu d'água é bastante comum em muitas partes da bacia do Amazonas, incluindo regiões do Brasil, por exemplo na área coberta pela Reserva de Desenvolvimento Sustentável Mamirauá. O apetite dos colombianos por esta criatura tornaram-na um bem relativamente escasso nesse país e a pesca atravessou fronteiras, para o lado brasileiro, essencialmente dedicada à exportação para a Colômbia. O efeito nos botos vermelhos tem sido devastador. Não se trata de problemas de golfinhos presos em redes, ou dos pescadores a tentarem livrar-se de um competidor indesejado. Não, é uma história de desperdício criminoso, revoltante, e mesmo com requintes de malvadez. [... ler mais]
Não sou muito de puxar para o sentimentalismo ou procurar reacções emotivas nas minhas contribuições, mas quando descobri o que se passava nas páginas do Projeto Boto devo confessar que senti uma imensa revolta. O que se segue é em grande parte emprestado das páginas do Projecto Boto, embora esta parte esteja apenas em inglês. O boto da imagem acima é um macho jovem que tem marcado o nome que os cientistas lhe deram, "Vani". Os investigadores do Projecto Boto aperceberam-se dele pela estranha barbatana dorsal e cicatrizes no corpo. Quando finalmente o capturaram ficaram chocados quando perceberam o que tinha sucedido ao animal. A aparência do animal é consequência de ter sido mutilado com uma faca. Além disso havia uma corda atada firmemente em torno da cauda do animal. Se não tivesse sido retirado o nó, que já estava a cortar a pele do animal, teria levado à queda da cauda, devido à interrupção do fluxo de sangue. Felizmente a corda foi cortada a tempo e o Vani foi visto um ano depois, recuperado das suas feridas. Não se trata apenas de um trabalho de um sádico sem propósito, os investigadores pensam que a corda teria sido colocada para segurar o boto até que fosse preciso para servir de isco. Isco para quê? Bem, os pescadores que trouxeram a pesca do urubu d'água para o Brasil trouxeram também o hábito repugnante de usar golfinhos e jacarés como isco. Esta caça é neste momento generalizada e o número de botos tem caído de forma acentuda. A espécie corre o risco de desaparecer da zona.
O boto da imagem acima é um macho jovem que tem marcado o nome que os cientistas lhe deram, "Vani". Os investigadores do Projecto Boto aperceberam-se dele pela estranha barbatana dorsal e cicatrizes no corpo. Quando finalmente o capturaram ficaram chocados quando perceberam o que tinha sucedido ao animal. A aparência do animal é consequência de ter sido mutilado com uma faca. Além disso havia uma corda atada firmemente em torno da cauda do animal. Se não tivesse sido retirado o nó, que já estava a cortar a pele do animal, teria levado à queda da cauda, devido à interrupção do fluxo de sangue. Felizmente a corda foi cortada a tempo e o Vani foi visto um ano depois, recuperado das suas feridas. Não se trata apenas de um trabalho de um sádico sem propósito, os investigadores pensam que a corda teria sido colocada para segurar o boto até que fosse preciso para servir de isco. Isco para quê? Bem, os pescadores que trouxeram a pesca do urubu d'água para o Brasil trouxeram também o hábito repugnante de usar golfinhos e jacarés como isco. Esta caça é neste momento generalizada e o número de botos tem caído de forma acentuda. A espécie corre o risco de desaparecer da zona.
Os requintes de crueldade dos pescadores de piracatinga vão no entanto ainda mais longe. Os investigadores do Projeto Boto têm encontrado crias atingidas por arpões, mutiladas e moribundas, algumas com cordas atadas em torno da cauda. Eis abaixo um exemplo de um pequenita que foi encontrada ainda com vida. Apesar dos esforços para a salvarem, esta cria morreu nos braços dos investigadores enquanto a mantinham à superfície da água e a tentavam levar para o laboratório para tratamento. Uma autópsia mostrou que mesmo que tivessem conseguido chegar um pouco antes teria sido já demasiado tarde. A pequenita tinha sido arpoada uns dias antes, e tinha uma infecção generalizada na região da cauda, com gangrena dos tecidos. A mãe desta pequena fêmea tinha permanecido perto dele até ao fim. Os investigadores estão convencidos que o sofrimento que a cria suportou durante esses dias terá sido infligido por um pescador no intuito de atrair a mãe e poder arpoá-la de perto. Este descrição perturbou-me, sobretudo porque ao procurar material sobre os botos tinham encontrado este filme de uma mãe a pertilhar um peixe com a sua cria.
Apesar dos esforços para a salvarem, esta cria morreu nos braços dos investigadores enquanto a mantinham à superfície da água e a tentavam levar para o laboratório para tratamento. Uma autópsia mostrou que mesmo que tivessem conseguido chegar um pouco antes teria sido já demasiado tarde. A pequenita tinha sido arpoada uns dias antes, e tinha uma infecção generalizada na região da cauda, com gangrena dos tecidos. A mãe desta pequena fêmea tinha permanecido perto dele até ao fim. Os investigadores estão convencidos que o sofrimento que a cria suportou durante esses dias terá sido infligido por um pescador no intuito de atrair a mãe e poder arpoá-la de perto. Este descrição perturbou-me, sobretudo porque ao procurar material sobre os botos tinham encontrado este filme de uma mãe a pertilhar um peixe com a sua cria.
Após procurar um pouco encontrei números do IBAMA. Entre Maio e Dezembro de 2001 cerca de 140 toneladas de piracatinga passaram pelo porto de Tabatinga em direcção à Colômbia. Este negócio envolvia na altura a matança de cerca de 8,000 jacarés e 150 botos por ano. Quanto aos jacarés não há perigo de extinção, são milhões, e recuperam rapidamente. Para os golfinhos, mesmo esquecendo os aspectos emotivos, este tipo de exploração é insustentável. Os botos vivem de 30 a 40 anos, mas crescem lentamente. As fêmeas só dão à luz a primeira cria por volta dos 6 a 8 anos de vida, e o intervalo entre crias é de 4 a 5 anos. É preciso parar a actividade antes que a espécie se extinga. O problema começou há poucos anos, tem vido a agravar-se, e é possível que por esta altura sejam mortos alguns milhares de animais por ano, incluindo dentro da própria reserva de Mamirauá. A caça ocorre em zonas inundadas, sendo por isso difícil de deter ou fiscalizar e, para além disso, os envolvidos usam a força e ameaças para silenciar eventuais testemunhas. Os investigadores do Projeto Boto estão no entanto optimistas, pois a quase totalidade da população local opõe-se fortemente a esta práctica, pelo que esperam que se consiga eventualmente acabar com ela. Espero bem que sim.
Para a semana voltarei aos botos, mas para falar de artigos científicos e não de aspectos tão macabros. A próxima contribuição será sobre as vocalizações destas fantásticas criaturas.
Nota: na pesquisa que fiz sobre o tema a piracatinga é o Calophysus macropterus, no entanto nas páginas do projecto boto fala-se do Cetopsis coecutiens. Se alguém me pudesse esclarecer de qual das criaturas se trata agradecia muito.
Ficha técnica
Foto do urubu d'água tirada por Mark Smith obtida por intermédio do PlanetCatfish.
Outras fotos e inspiração para o texto provenientes das páginas do Projeto Boto.

